
肺鱼特征整理:修订间差异
自动添加 Sofia 模板和分类 |
|||
| 第145行: | 第145行: | ||
■类似于硬骨鱼的特征:肩带有锁骨和匙骨的加入;有膜原骨的出现;鳔有内褶,类似于总鳍鱼。 | ■类似于硬骨鱼的特征:肩带有锁骨和匙骨的加入;有膜原骨的出现;鳔有内褶,类似于总鳍鱼。 | ||
{{学科分类}} | |||
[[Category:动物学]] | |||
2025年8月23日 (六) 16:31的最新版本
肺鱼亚纲共有特征
- 颌关节自接型,与陆生脊椎动物相似。
- 具有高度特化适合压碎无脊椎动物甲壳的齿板。
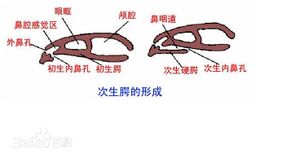
次生腭的形成 - 大部分骨骼为软骨;没有椎体,只有椎弓和脉弓,终生保留发达的脊索。(好逊...软骨鱼都完全退化了,其他硬骨鱼也都至少是完全骨化了,废鱼<・)))><<实至名归)
- 肩带有膜原骨加入(与软骨鱼不同),具有了锁骨和匙骨(倾向于硬骨鱼的特点)。
- 肠道内具有螺旋瓣,倾向于软骨鱼。
- 鳔向内褶皱,有鳔管,与食管相通,具有呼吸功能,与总鳍鱼类似。
- 不完全的两心房两心室,却具有动脉圆锥,静脉窦也有(注意心房和心室的间隔都不完全)。由于用肺呼吸,鳔动脉从第六对动脉弓的出鳃动脉发出,与脊椎动物肺动脉同源,而一般鱼类的鳔动脉是从背大动脉分支的。鳔静脉与肺静脉同源,直接返回心房左侧。
- 完全没有血管囊(甚至超越两栖类的进化;两栖类为退化的血管囊)。
- 神经系统较为发达,大脑发达而小脑不发达,侧脑室独立(再一次超越鱼类),脑皮有神经组织而不是纯上皮组织。
- 雌性输卵管更近似软骨鱼,有一对,发生上属于牟勒氏管。喇叭口靠前,接近心脏。两管末端合并,开口于生殖乳突顶,再通入泄殖腔。
- 和圆口类一样,肺鱼没有脾脏。[1]
- 具有罗伦氏壶腹。
- 没有次生颌(杨安峰《脊椎动物学》)。
- 与有尾两栖类相似,具有声门、会厌软骨。
- 脑下垂体的结构(出现神经部)及其分泌的激素、晶体蛋白、胆汁盐、鳃弓肌肉均与有尾两栖类相似。(杨安峰《脊椎动物学》)
- 不仅具有后主静脉 还具有后腔静脉 这两者并非同源结构 所以能够共存很合理
- 多被圆鳞

肺鱼亚纲的各个种类
单鳔肺鱼目
只有澳洲肺鱼一科一种。分布在澳大利亚昆士兰州(东南部)淡水。
澳洲肺鱼科
只有澳洲肺鱼一种,只有一个鳔代表着原始性;鳔管开口于腹部但鳔扭转到背面。体型侧扁,胸腹鳍粗壮,桡状,鳞大,相等的角质齿板,双列型鳍骨,以基鳍骨为中轴骨,辐鳍骨在两侧。成体没有休眠现象,幼鱼无外鳃。
双鳔肺鱼目
有非洲和美洲肺鱼。 鳗形鱼体,胸鳍较为狭短或鞭状,鳍上只有中轴骨,不是双列型。鱼鳞小,埋在皮下,有休眠(形成茧,只用鳔呼吸,是水分缺乏导致的现象),幼鱼有外鳃。
美洲肺鱼科
分布在南美洲淡水。只有一种。
5对鳃弓,偶鳍小而短,奇鳍低矮不发达,幼鱼外鳃存在时期短。
非洲肺鱼科
分布在非洲中部淡水,有三种。
6对鳃弓,偶鳍鞭状,奇鳍高,整个幼鱼期都有外鳃。
[2]只有两对鳃[1]
后人的一点补充
在愿程某文字解析中亦有部分关于肺鱼的精妙论述,有感于埋没实在可惜,故一同贴于此处。(若侵权请直接删除,感谢理解)
1.呼吸系统
常考知识点:
◆肺鱼可以通过肺直接呼吸空气,其肺内具有褶皱能够增大气体交换面积
拓展知识点:
◆澳洲肺鱼Neoceratodus为单鳔目,而非洲肺鱼Protopterus和美洲(南美)肺鱼Lepidosiren则合称为双鳔目Lepidosireniformes。澳洲肺鱼通常被认为是更原始的物种,其甲状腺具有功能上的缺陷因此被认为具有幼态持续现象(即未性成熟的幼体便具有繁殖能力),而其他两种肺鱼是否具有幼态持续现象则仍在研究当中。同时其还具有特殊的电感受器能够定位猎物。
◆南美肺鱼(美洲肺鱼)和非洲肺鱼具有发达的肺和较为退化的鳃,而澳洲肺鱼的肺则较为简单,其大部分情况下还是用鳃呼吸的。非洲肺鱼和美洲肺鱼的肺分为两叶,可称为双鳔类,而澳洲肺鱼只有右侧肺发育,左侧肺退化,因此可称为单鳔目,其单独的右侧肺同时由两根肺动脉支配。类似于陆生脊椎动物,前两者的中枢神经系统中具有CO2与氢离子的感受器,可以通过调节肺通气改变体液的酸碱状态,而关于澳洲肺鱼的相关信息则较少。在陆生脊椎脊椎动物中,中枢感受器对血液中二氧化碳浓度的调控占总体的70-80%,而外周感受器占20-30%。在美洲肺鱼种,外周感受器的调控水平与陆生脊椎动物相似(占高碳酸血症诱导的通气增加反应的20%),这表明中央和外周CO2/H+受体的起源非常早。在陆生四足动物和肺鱼中通过检测氧分压感知血液含氧程度(在美洲肺鱼中,中枢感受器位于第四脑室内)。同时非洲肺鱼P.aethiopicus具有肺牵张反射。
◆肺鱼的现存近亲矛尾鱼(拉蒂迈鱼coelacanth(Actinistia))同样具有一个肺,但是其内被脂肪填充
◆非洲肺鱼和美洲肺鱼会产生类似于两栖动物的表面活性剂,而澳洲肺鱼的所谓“肺泡表面活性剂”的成分则更类似于辐鳍鱼类,此外有证据表明澳洲肺鱼在长达3亿年的时间里一直保留着相同的表面活性剂组成。
◆肺鱼保留了第2到第6对动脉弓,其中第三对和第四对动脉弓在非洲肺鱼和美洲肺鱼只有连通背腹动脉的功能,第二对和第三对动脉弓所属的鳃退化,第四动脉弓所属的鳃也产生了功能性退化,只有第五对和第六对动脉弓能够将缺氧血送入鳃中进行气体交换。而澳洲肺鱼的II-V对动脉弓则能够直接将血液送到到未退化的有功能的鳃中。同时肺鱼还存在第六对动脉弓起源的肺动脉。下为非洲肺鱼动脉弓及鳃的模式图。
2. 循环系统
常考知识点:
■心脏具有不完全的两心室和两心房的分隔(实际上其具有两个分开的左右心房和一个合并的心室,其内具有瓣膜将单独的心室分为左心室腔和右心室腔,且缺少房室瓣),且具有不完全的双循环系统,同时动脉圆锥具有隔膜可以帮助动静脉血分离。

■鳔动脉从第六对动脉弓的出鳃动脉发出,与脊椎动物肺动脉同源,而一般鱼类的鳔动脉是从背大动脉发出的
拓展知识点:
◆在水中进行鳃呼吸和陆地上进行空气呼吸时具有不同的血流途径:在水中(FW)进行鳃呼吸时,其肺动脉部分收缩,使得大部分血液经鳃进行气体交换后直接进入背大动脉并给全身组织供血;而当进行夏眠(AE)时,肺动脉开放且鳃片坍缩,大部分血液不经过鳃而直接进入肺,并重新回到心脏中,通过心脏泵送到全身组织中(下图为上述过程的模式图)。
◆肺鱼的心脏结构
肺鱼心脏已经分化出了独立的左右心室,左心室容纳肺静脉来源的动脉血而右心室容纳静脉窦来源的静脉血。在其他硬骨鱼中动脉血经过鳃交换氧气后直接进入背大动脉供给全身组织而不重新经过心脏而在肺鱼中血液经过肺进行气体交换后经肺静脉重新回到心脏中。同时肺鱼的心室和心房也很特殊
其心室和心房之间没有房室瓣,因此心房和心室统称心房室atrioventricular(简写为AV)。其心脏内起到房室瓣作用的则是一个具有软骨核心的球状塞,称为房室塞atrioventricular plug,这个塞子的背侧壁与静脉窦融合,而其它面则游离。肺鱼的肺静脉也十分特殊,其直接穿入静脉窦中,并贴着静脉窦腹侧行进(见下图1)。
Fig1.SV为静脉窦,LA为左心室,RA为右心室,白色箭头所指的即为肺静脉
随后肺静脉继续上行,到达房室塞后会碰到一个复杂的褶皱,称为肺静脉褶pulmonalis fold(见图2)肺静脉的壁随后与静脉褶融合,最终到达左心房。因此由肺静脉-肺静脉褶构成的一个通道能够将肺静脉中的动脉血与静脉窦中的静脉血相分离,从而在进入心脏之前就形成了两者的分离,类似于陆生的哺乳动物肺静脉将动脉血输送进入左心房,而静脉窦(或其衍生结构)将静脉血输送至右心房。
Fig2.P为房室塞,SV为静脉窦,PV为肺静脉,F为肺静脉褶,LA为左心房,RA为右心房。


Fig³.RV为右心室腔,LV为左心室腔,P为房室塞,白色箭头指的是血流出心脏的出口,后接动脉圆锥+动脉球

Fig4.为处于不同状态下的肺鱼心脏横切,A为心房,V为心室,星号指的是房室塞内的软骨核心。ab图为非夏眠状态下,而c为夏眠状态,可见箭头所指的动脉褶大小发生明显减小,而其他的部分则无显著的组织学与结构的变化。
Fig3图中可以看见发达的心室隔膜,将心室分为左右两个心室腔,但仍然分隔不完全。
Fig4可见肺鱼接收心脏泵血的部分:血液流出道OFT非常长,其从房室塞处一直延伸到鳃部的鳃动脉分支处。其在体内的行进路线较为复杂,具有两个弯曲点,将整个OFT分为三个部分:远端、中段和近端部分。从结构上看,近端部分与其他鱼类的动脉圆锥同源,而中段和远端部分与动脉球同源。动脉圆锥具有发达的,血管化良好的心肌壁,并且具有圆锥瓣膜(conus valves)。在非洲肺鱼Protopterus中,这些瓣膜位于OFT的腹侧和背侧壁上排列成两到三排。值得注意的是,大部分非洲肺鱼的动脉圆锥瓣膜退化成了无瓣膜功能的横突,其似乎在心脏血液分流的方面不起重要作用。动脉球则的壁与动脉类似,由弹性纤维和胶原纤维组成,同时表面覆盖了一层薄薄的心肌,与动脉圆锥的心肌层相连。
肺鱼的OFT的一个显著特征即具有两个松散的组织,称为动脉球褶,其中较为明显的一个称为螺旋褶,其从近端OFT一直延伸到远端ORF;另一个则称为动脉球褶,其从中段OFT延伸至远端OFT。这两个瓣膜具有相似的结构,其被心内膜所包裹并且仅具有少量的胶原蛋白和弹性蛋白,以及具有少数分散的成纤维样细胞分布。这两个褶皱能够形成不完整的螺旋状分隔,有人推测,这些褶皱有助于在心脏的这一区域将血液流分开,形成腹侧和背侧血流。肺鱼缺乏腹部主动脉,其三对鳃动脉直接从OFT处发出。
由于不完整的分流机制,含氧血和缺氧血在心脏中仍会发生混合,含氧血优先进入背主动脉,而缺氧血优先流入肺动脉。早期研究表明,当向肺静脉注射不透明介质时,显影剂会充满左心室、OFT的腹侧通道以及腹侧的鳃弓。相反,当造影剂注入静脉窦时,它会充满右心室、OFT的背侧通道以及后面的几个鳃弓。值得注意的是,前面的鳃弓中没有鳃丝(肺鱼只有后面两对鳃弓就有鳃丝,前面的三对均无鳃丝)。前面的鳃弓是向脑部和心脏供血血管的起源部位,也是体动脉(背主动脉)的起源部位。前鳃弓中没有鳃丝可以最大限度地减少氧气向静止、缺氧水中扩散所产生损失。后鳃弓中有发育良好的鳃丝,肺动脉从后部的几个鳃弓发出。背主动脉和肺动脉之间仍然通过管道或血管保持连通。同时在背主动脉和肺动脉的发出基部具有可以收缩的血管成分,其能够调控血液在体循环和肺循环中的分布。当处于低氧环境时,更多的血液会分配到肺循环中。这些可以舒缩的血管节段具有增厚的血管壁中间层,同时具有胞质内含颗粒的细胞,并且其受到迷走神经支配。因此该阶段能够受内分泌和神经系统的调
节。
◆夏眠时肺鱼的心脏及循环系统的结构变化
当肺鱼处于夏眠时,动脉球褶会缩小从而使其在OFT内排列的更加规整(见Fig4),使得血液分流更加完全和明显,这可能是因为夏眠时脱水所导致的。这种变化能够最小化血液在OFT中的混合。尽管在解剖学上未发生显著变化,其在单细胞水平和亚细胞水平上仍具有显著变化。在非休眠状态下,电镜照片中显示出其胞质内具有大量的电子密度较低的物质填充,而在休眠状态下则缺少这种低电子密度物质(见Fig5)。随后通过PAS染色证明,这种低电子密度物质即为糖原,其能够在夏眠期间为心肌细胞提供能量。

Fig5.ab为非夏眠状态下的心肌细胞,a图中箭头指的是胞质中含有的低电子密度物质,而b图中星号则为箭头所指部分放大;cd图为夏眠状态下的心肌细胞,可见星号部分缺失了b图所示的低电子密度物质。
夏眠还伴随着心肌细胞核中染色质分布模式的变化。非夏眠状态下的肺鱼大多数心肌细胞显示出典型的异染色质模式,这是低转录、低能量状态的典型特征。相反,与人们可能预期的不同的是,大多数心肌细胞在夏眠期间显示出常染色质模式,这可能反映了代谢活动的增加。尽管这一特征的真正意义尚不清楚,但夏眠不能被视为简单的功能和代谢活动(包括基因表达)的限制。
除了心脏结构和功能的改变外,夏眠中还将鳃与环境隔离并建立专有的肺呼吸过程。此时鳃被一层保护性黏液包裹,同时部分鳃片坍缩。因此肺成为机体的唯一氧气供应源,此时血流经过血管基部的舒缩部分调控发生重分配,大部分的血液进入肺动脉而非鳃弓动脉。同时连接入鳃动脉和出鳃动脉的旁路血管打开,使血液直接绕过鳃循环。值得注意的是,在夏眠阶段肺鱼的肺会发生显著的改变,其血管分布更加丰富,并且更加膨大,内部的嵴和褶更加明显,表现为肺的生理性膨大
◆肺鱼可供记忆的特殊点:
■肺鱼的独特特征:有高度特化的用于压碎无脊椎动物甲壳的齿板;非洲肺鱼具有六对鳃弓,且完全依靠肺呼吸;现生肺鱼有许多幼鱼的特征,暗示了幼态成熟的可能;具有眼睑和瞬膜;心脏不完全两心室两心房,具有不完全的双循环系统,同时动脉圆锥具有隔膜可以帮助动静脉血的分离;自接型;没有次生腭但是有内鼻孔;侧脑室独立;完全不具有血管囊;没有椎体,只有椎弓和脉弓,终生保留发达的脊索;由于用肺呼吸,鳔动脉从第六对动脉弓的出鳃动脉发出,与脊椎动物肺动脉同源,而一般鱼类的鳔动脉是从背大动脉分支的;泄殖腔膀胱。
■类似于软骨鱼的特征:大部分骨骼终身软骨,仅顶部具有少量膜原骨;生殖管发育类似于软骨鱼;具有动脉圆锥和螺旋肠瓣;大脑皮层具有神经细胞。
■类似于硬骨鱼的特征:肩带有锁骨和匙骨的加入;有膜原骨的出现;鳔有内褶,类似于总鳍鱼。