
脊比笔记:循环系统:修订间差异
| 第57行: | 第57行: | ||
*辐鳍鱼的侧腹静脉通常消失,腹鳍由后主静脉引流。来自鳔的血液排入肝静脉或总主静脉。 | *辐鳍鱼的侧腹静脉通常消失,腹鳍由后主静脉引流。来自鳔的血液排入肝静脉或总主静脉。 | ||
*肺鱼的静脉回流到心脏与其他鱼类的相似,只是右侧后主静脉会扩大,承担从身体后部引流血液的大部分责任,因此通常称为后腔静脉(图12. | *肺鱼的静脉回流到心脏与其他鱼类的相似,只是右侧后主静脉会扩大,承担从身体后部引流血液的大部分责任,因此通常称为后腔静脉(图12.23c)。成对的侧腹静脉融合形成不成对的腹侧腹静脉,该静脉将腹鳍中的血液引流并排入静脉窦。肺鱼也有单一的腹静脉。 | ||
*两栖类 | *两栖类 | ||
2025年1月13日 (一) 20:54的版本

主动脉弓
- 原始主动脉弓和它们穿过的鳃弓的数量仍存在争议。一些甲胄鱼类有多达10对鳃弓和大概10对主动脉弓。
- 主动脉弓对的数量在生物体中有所不同。七鳃鳗有8对(图12.12),盲鳗有15对。一些鲨鱼种类拥有10或12对。
- 在大多数有颌鱼和所有四足动物的胚胎发育过程中,通常只出现最多六对。
- 软骨鱼
- 软骨鱼的第一鳃裂变小为喷水孔。第一鳃裂的动脉弓腹侧部分未出现,入鳃血流由相邻的收集环提供。第一动脉弓背侧部分将这些含氧的血流导出喷水孔。
- 这些收集环是额外增加的部分,不是原始性状。
- 从胚胎学上来说,颈外动脉起源于腹主动脉的前端,它后来与收集环相连,将含氧血液输送到下颌。这是与其功能相适应而发生的变化。
- 故曰:软骨鱼五入四出,是Ⅰ-Ⅴ入,Ⅱ-Ⅴ出。Ⅰ出改道别的地方去了,不会通入背主动脉。
- 硬骨鱼
- 大多数辐鳍鱼是四入四出,喷水孔减小或消失,第一动脉弓完全消失了。
- 澳洲肺鱼:Ⅲ-Ⅵ四个弓供应五个鳃裂
- 非洲肺鱼:第三四个鳃消失,但Ⅲ、Ⅳ弓仍存在,其间无毛细血管。
- 三种肺鱼都用Ⅵ发出通向鳔的动脉,但Ⅵ没有失去和背主动脉的联系。
- 人们曾经认为Ⅲ Ⅳ两弓没有鳃,会把缺氧血液直接导入背主动脉,导致呼吸效率降低。实际上并不会,后面详述。
- 两栖类:前两个主动脉弓(I、II)在发育早期消失。
- 大多数有尾目幼体中,三个主动脉弓(III-V)带有外鳃,最后一个主动脉弓(VI)会长出通向正在发育的肺的肺动脉。
- 例外是Necturus泥螈,Ⅵ的一部分消失,只有背部部分存留下来,形成肺动脉的基部。
- III和IV之间的背主动脉称为颈动脉导管(carotId duct),通常在变态时关闭。这迫使颈动脉充满来自腹主动脉的血液。
- III和IV之间的腹主动脉弓部分成为颈总动脉(common carotId artery),为外颈动脉(来自前部的腹主动脉)和内颈动脉(背主动脉前部和第三主动脉弓)提供血液。
- IV、V构成连接背主动脉的主要全身血管。
- VI也连接背主动脉,其最后一段形成动脉导管。在汇入背主动脉前不久,第六主动脉弓分出肺动脉,肺动脉又分成小分支,通向口底、咽部和食道,然后进入肺部。
- 无肺有尾目的肺动脉(若其存在)则为颈部和背部的皮肤供血。
- 无尾目幼体通常由四个内鳃(Ⅲ-Ⅵ)。肺动脉从Ⅵ发芽.
- 变态时,这些鳃与颈动脉导管和主动脉弓V一起消失。
- 弓VI失去了与背主动脉的连接,成为肺皮动脉,一个分支是肺动脉,另一个是皮动脉,它沿着背部和侧面的体壁将血液输送到皮肤。
- 大多数有尾目幼体中,三个主动脉弓(III-V)带有外鳃,最后一个主动脉弓(VI)会长出通向正在发育的肺的肺动脉。
- 爬行类
- 最显著的解剖学改变是腹主动脉的分裂。在胚胎发育过程中,腹主动脉分裂形成离开心脏的三条独立动脉的基部:左主动脉弓、右主动脉弓和肺动脉干。
- 右体系统弓往往是两者中更突出的一个,因为它供应额外的血管。例如,在较原始的脊椎动物中,颈动脉起源于腹主动脉,而在爬行动物中则起源于右体弓。
- 在大多数爬行动物中,锁骨下动脉从背主动脉分支出来,但在某些爬行动物中,它们从体系统弓分支出来。
- 鸟类
- 通向翅膀的成对的锁骨下动脉来自颈内动脉而不是背主动脉
- 两条颈总动脉可以分别从右侧体弓发出,也可以两者合并形成一条颈动脉
- 头臂动脉是一种短小但重要的血管,存在于少数爬行动物(尤其是乌龟)中,但在许多鸟类中却是主要的前部血管。
- 哺乳动物
- 左锁骨下动脉从左体动脉弓分出,而右锁骨下动脉包括右主动脉弓(IV)、相邻的右背主动脉的一部分,以及从这些部分长入右肢的动脉
- 两条颈总动脉可能由同一条头臂动脉分支而来,也可能在主动脉弓上有分别的起源。
静脉系统
在发育早期,存在三组成对静脉:来自卵黄囊的卵黄静脉(vitelline veins)、来自胚胎本身身体的主静脉(cardinal veins)和来自骨盆区域的侧腹静脉(lateral abdominal veins)。
- 卵黄静脉最早出现,发生在卵黄的上方。这两条静脉通过卵黄柄进入胚胎,向前生长连接到静脉窦.肝脏的原基发育过程中侵入了卵黄静脉,在卵黄静脉内部发育,卵黄静脉的内墙即被分隔为肝窦,肝脏和静脉窦之间的卵黄静脉就被称为肝静脉.
- 主静脉包括两条前主静脉和两条后主静脉,在心脏水平回合为总主静脉,汇入静脉窦.
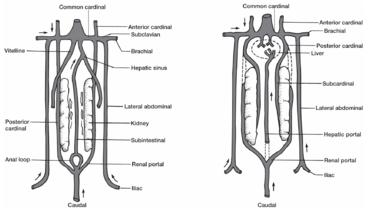
- 侧腹静脉收集来自后肢/腹鳍的血液.
- 鱼类:侧腹静脉→髂静脉→锁骨下静脉←肱静脉
- 四足动物:侧腹静脉→肝脏;锁骨下静脉→心脏
- 两栖类:侧腹静脉融合为一条腹静脉。
- 在鳄鱼、鸟类和哺乳动物中,侧腹静脉不存在。
- 肝门静脉:由胚胎的肠下静脉发育而来,从尾静脉发出的不成对的血管。肠下静脉在肛门周围形成一个环并向前延伸,沿着肠道腹壁走行,从肠道收集血液。它从肠道穿过肝脏,最后汇入左卵黄静脉。在增生的肝索中,卵黄静脉分解成小的肝窦网络。肠下静脉的前端将血液排入这些肝窦,而其后端则退化并失去与尾静脉的联系,称为肝门静脉。它从肠道、胃、胰腺和脾脏收集血液。
- 肾门静脉:发育早期,尾静脉的血液流经肠下静脉或后主静脉,而后主静脉是更常见的路径。后主静脉延伸至肾脏,向前流入心脏。随后,一组下主静脉出现在肾脏之下,接受肾脏血液,向前流入后主静脉。下主静脉建立起来,下主静脉和肾脏的连接处之间的后主静脉就会萎缩。在发育的这个阶段,肠下静脉也失去了与尾静脉的连接。由于这些血管改变,尾部的血液现在必须流经肾脏。随着尾部血液通过肾脏,尾静脉成为肾门静脉系统。血液从肾脏通过新建立的下主静脉排出(图12.21b)。
- 肺静脉:肺静脉不是通过现有血管通道的变形而产生的。相反,许多小血管在胚胎肺芽内分别起源并排出胚胎肺芽。然后它们汇聚成几条公共血管,成为进入左心房的肺静脉。

各个类群的静脉系统
- 辐鳍鱼的侧腹静脉通常消失,腹鳍由后主静脉引流。来自鳔的血液排入肝静脉或总主静脉。
- 肺鱼的静脉回流到心脏与其他鱼类的相似,只是右侧后主静脉会扩大,承担从身体后部引流血液的大部分责任,因此通常称为后腔静脉(图12.23c)。成对的侧腹静脉融合形成不成对的腹侧腹静脉,该静脉将腹鳍中的血液引流并排入静脉窦。肺鱼也有单一的腹静脉。
- 两栖类