跳转到内容
(
差异)
←上一版本 | 最后版本 (差异) | 下一版本→ (差异)
总论
文件:676.png图1:苔藓植物的进化地位
- 苔藓植物属于孢子植物,是最低等的有胚植物。
- 像所有有胚植物那样,苔藓植物有二世代交替,属于居间减数分裂,但有胚植物中仅有它是配子体占优势。(旧时有观点认为苔藓植物是植物进化中的盲端,此学说现已被抛弃,苔藓植物的进化地位见图1)
- 苔藓植物一般分三类,按进化顺序排:苔(俗称Liverwort,学名Hepaticae)、藓(俗称Mosses,学名Musci)、角苔(俗称Hornwort,学名Anthocerotae)。
- 苔藓植物没有根、茎、叶的分化,只有假根和叶状体(或具叶体),没有维管,所以长不高。
- 每个苔藓植物个体(配子体)由单倍体的孢子发育而来,先形成原丝体(Protonema),然后形成叶状体,最后长出配子囊,通过有丝分裂产生配子,借助水使雌雄配子相遇,产生附着在配子体上的孢子体,它成熟后减数分裂产生孢子。
- 苔藓植物的生殖离不开水,只能生长在潮湿的地方。
苔
文件:677.png图2:地钱纲模式图
文件:681.png图3A:地钱纲
文件:678.png图3B:地钱纲
文件:682.png图4A:地钱纲的营养组织
文件:683.png图4B:地钱纲的生殖组织
文件:684.png图5:叶苔纲
 图6:叶苔纲的组织
图6:叶苔纲的组织
- 苔的共同特征是:细胞具有油体(Oil Body)、孢子囊中有弹丝(Elater)。
- 油体是萜类物质的液滴,有一层膜包被。
- 苔的配子体有两种:叶状的(Thalloid)和具叶的(Foliose)。
- 苔分两纲:地钱纲(Marchantiopsida)、叶苔纲(Jungermanniopsida),具叶型只出现于后者。(但后者也包含很多过渡形态的物种)
- 地钱纲:
- 配子体为叶状体,是一团扁平的铺在地面的组织,有时呈二叉分支。
- 叶状体底面向下伸出假根(Rhizoid),它是背光生长的,每根假根是一个细胞,不分支。(也有多细胞组成的Venter Scale,见图4A)
- 假根有固着和吸水的功能。
- 叶状体上表面有一层防水的角质层。
- 上表面细胞间隙的部分区域下陷形成气室(Air Chamber),既能交换气体也能促进吸水。
- 气室的底部是一群叶绿体丰富的薄壁细胞,是同化组织。
- 气室和维管植物的气孔在进化上无关,它的入口由一个或多个环状细胞控制。
- 叶状体下部的细胞,叶绿体较少,主要起贮藏作用,油体集中于这些细胞。
- 叶状体有中脉(Mid Rib,见图3A),从中脉上长出芽孢杯(Gemma Cup),具有无性生殖功能,在有水滴进入时发挥功能。
- 从配子体向上伸出雌生殖托(Archegoniophore)和雄生殖托(Antheridiophore)。(图3B)
- 地钱是雌雄异体,配子体的性别由单条性染色体决定。
- 雌配子囊在雌生殖托底面,向下有狭长开口用于接受雄配子。
- 雌配子囊以辐射状排列,每条辐线有总苞(Involucrum)保护。
- 雌配子囊母细胞首先分裂成四个细胞,周围三个Jacket Cell环绕着中心的Axial Cell。
- Jacket Cell将来发育成雌配子囊壁的中间部分。(底部由雌配子囊母细胞下方的Stalk Initial发育而成)
- Axial Cell首先分裂成Cover Cell(下部)和Inner Cell(上部)。
- Cover Cell起保护作用,Inner Cell经多次分裂,在顶上形成1个细胞,周围形成4-8个细胞,中间形成卵细胞。
- 雄配子囊在雄生殖托上面,向上开口用于释放雄配子。
- 雄配子囊母细胞首先分裂成4个细胞,呈十字。
- 然后每个细胞分裂成靠内和靠外的两个细胞。
- 外部的细胞发育成雄配子囊壁,内部的细胞发育成雄配子。
- 受精发生于雨天,雨水将雄配子带至地面,当雨滴溅起水花时把它带入雌配子囊,发育成附着在配子体上的孢子体。
- 地钱的孢子体呈椭球形,没有杆或只有很短的杆。
- 孢子体几乎没有光合作用的功能。
- 孢子体的壁只有一层细胞,在尖端有两层。
- 最初,孢子体有雌配子囊壁(称为Embryotheca,胚鞘)包被,后破裂,在基部呈鞘状。
- 孢子体中心的细胞每次分裂形成一个孢子母细胞和一个弹丝细胞,前者减数分裂形成4个孢子,后者不分裂。
- 一般地,孢子和弹丝细胞的个数比为4:1,在个别物种中可变为8:1甚至128:1。
- 成熟时,孢子体尖端破裂,一部分细胞凋亡,另一部分则向上卷曲呈牙状。
- 叶苔纲:
- 叶苔纲没有气室,没有生殖托。
- 油体广泛分布于所有细胞,不是集中于贮藏细胞。
- 叶状体有中脉,无Venter Scale。
- 具叶体由杆和三列叶片组成,上侧两列较大,下侧一列较小或无,叶没有中脉。(它的叶和维管植物的叶在进化上无关)
- 芽孢杯生在叶的尖端和边缘,较小。
- 部分物种中,雌配子囊生在叶状体的背侧。(一般物种的雌配子囊都源于叶状体顶端细胞,而它们的源于顶端细胞下方的一个细胞)
- 孢子体有较长的杆将它抬高,适应于利用风传播孢子。
- 孢子体成熟时,整个孢子体壁呈四瓣裂开。
藓
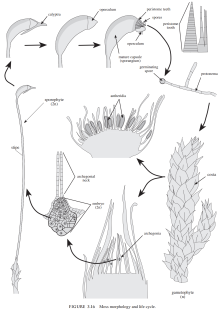 图7:藓的模式图
文件:680.png图8A:藓
文件:686.png图8B:泥煤藓
文件:687.png图9:泥煤藓和黑藓
文件:688.png图10:藓纲
文件:689.png图11:藓的配子囊发育
图7:藓的模式图
文件:680.png图8A:藓
文件:686.png图8B:泥煤藓
文件:687.png图9:泥煤藓和黑藓
文件:688.png图10:藓纲
文件:689.png图11:藓的配子囊发育
- 藓是苔藓植物中物种最丰富的类群。
- 从藓开始出现真正的气孔,藓的气孔由两个保卫细胞组成。
- 藓的孢子有周壁层,这可能也是藓和维管植物的共同特征之一。
- 藓的配子体形态类似苔的具叶体。
- 藓的孢子体有很长的杆(Seta)将它抬高,这结构将来进化为维管植物的茎。(泥煤藓纲和黑藓纲则没有或很短,这些物种的孢子体有源自配子体的伪足(Pseudopodium)将它抬高)
- 部分藓的物种出现了输导组织,Hydroid负责输水,Leptoid负责输送糖分,一般认为这些结构和维管植物的维管束无关。
- 横切面上,几乎总是Hydroid在里面,Leptoid在外面,Hydroid细胞边上有Stereid细胞(是活细胞,有细胞核,细胞壁加厚),Leptoid细胞边上有薄壁细胞(它的细胞壁没有胞间连丝)。
- 藓的配子体分为三部分:假根(Rhizoid)、假茎(Cauloid)、叶状体(Phylloid)。
- 每根假根由多细胞组成,可分枝,这与苔不同。
- 藓分为三纲:泥煤藓纲(Sphagnopsida)、黑藓纲(Andreaopsida)、藓纲(Bryopsida)。
- 泥煤藓纲:(黑藓纲与之类似)
- 泥煤藓是泥煤的最主要来源,是经济重要物种。
- 泥煤藓生活在酸性的沼泽中,像地毯一样铺在地上。
- 泥煤藓的细胞壁有木质素。
- 泥煤藓的孢子在某种真菌的刺激下发育,长出直立的假茎,并在间隔均匀的地方分枝,每年顶端停止生长而离顶端最近的分枝发育为主干,形成假二叉分枝形态。
- 底部的假茎和分枝会逐步死亡变成泥煤,此时附着其上的活着的分枝就成了独立的植株,可视为一种营养繁殖。
- 泥煤藓的假茎的表皮是几层死细胞,能通过毛细作用吸水。
- 在显微镜下,可看到泥煤藓的“叶”的结构为许多小的含叶绿体的同化细胞围绕着几个大的用于储水的死细胞。
- 雄配子囊长在叶的尖端,有长杆,而雌配子囊长在枝的尖端,无长杆。
- 精卵结合后长出孢子体,其杆很短,但雌配子囊长出伪足。
- 最初孢子体有胚鞘(雌配子囊壁)包被,后破裂,一部分组织留在孢子体尖端,称为蒴帽(Calyptra),注意它是单倍体的。
- 随着孢子体内部气压增大,孢子体尖端破裂,能听见声音,蒴帽凋亡。
- 藓纲:
- 藓纲的孢子首先发育成绿丝体(Chloronema),它的分枝很多,细胞富含叶绿体,且细胞之间的隔膜垂直于主轴。
- 末端的绿丝体长成轴丝体(Caulonema,又称茎丝体),它的细胞叶绿体很少,细胞之间的隔膜是斜着的。
- 在光照充足的条件下,轴丝体长出藓的个体。
- 配子囊长在枝的末端,枝梢既可只有单性配子囊也可有双性配子囊,个体既可雌雄同体也可雌雄异体。
- 配子囊的发育见图11。
- 精卵结合后,孢子体长出很长的杆(称为蒴柄,Seta),将它抬高。
- 蒴柄上部称为朔台(Apophysis),此处气孔丰富。
- 孢子体尖端也有蒴帽。
- 孢子体分外层组织(Amphithecium,又称朔周层)和内层组织(Endothecium)。
- 外层组织起保护作用。
- 内层组织的最外层产生孢原细胞,分裂形成孢子母细胞,再减数分裂形成孢子。
- 内层组织的内部组织不育,称为朔轴(Columella),功能是提供营养。
- 朔轴通过吸根(Haustorium)伸入配子体中吸收水分和糖类。(但孢子体和配子体之间没有胞间连丝)
- 孢子体尖端有朔盖(Operculum),朔盖的边缘有环带(Annulus),环带的细胞能分泌黏液,吸水时膨胀帮助朔盖破裂。
- 朔盖的下方有朔齿(Peristome)。
角苔
文件:691.png图12:角苔
文件:690.png图12:角苔的组织
- 角苔的化石证据很少,所以较难确定它的进化地位,但它具有大多数藓和维管植物共有的特征,因此一般将它放在藓和维管植物之间。
- 角苔的每个细胞只有一个叶绿体,它富含淀粉核。
- 配子体和孢子体都有气孔。
- 孢子体出现了居间分生组织,可以无限生长,为孢子体独立做好了准备。
- 孢子体附着在配子体的方式已接近松叶蕨(Psilophytina),不同于其它苔藓植物。
- 孢子体从配子体向上长高,能进行光合作用。
- 居间分生组织在孢子体基部,周围有配子体形成的鞘保护。
- 孢子体只有最底部和配子体相连,此处称为足(Foot)。
- 孢子体的中心是几列不育细胞,用于输送营养,周围是大量的孢原组织,最外层是孢子囊壁。
- 孢子体中有类似弹丝的细胞,但来源不同,故称为伪弹丝(Pseudo-elater)。

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}