
第二章 原生质:质膜,细胞核,细胞器
细胞是生命的最小结构和功能单位(Sitte, 1992)。生物体由单个细胞或细胞复合体组成。细胞在大小、形态、结构和功能上差异很大。有些细胞的尺寸以微米计,有些以毫米计,还有一些以厘米计(某些植物中的纤维)。有些细胞执行多种功能,而另一些则在其活动中具有专门化。尽管细胞之间存在极大的多样性,但它们在物理组织和生化特性上却非常相似。
细胞是生物结构和功能的基本单位这一概念基于细胞理论(cell theory),该理论由Mathias Schleiden和Theodor Schwann在19世纪上半叶提出。1838年,Schleiden得出结论,所有植物组织都由细胞组成。一年后,Schwann(1839)将Schleiden的观察扩展到动物组织,并提出了所有生命的细胞基础。1858年,当Rudolf Virchow概括出所有细胞仅来自已有细胞时,所有生物体由一个或多个细胞组成的概念具有了更广泛的意义。在其经典形式中,细胞理论提出,所有植物和动物的身体都是个体分化细胞的集合体,整个植物和动物的活动可以被视为个体组成细胞活动的总和,其中个体细胞至关重要。
到19世纪下半叶,提出了细胞理论的替代理论。被称为有机体理论(organismal theory),它认为整个有机体不仅仅是一组独立单位,而是一个被细分为细胞的活体单位,这些细胞相互连接并协调成一个和谐的整体。常被引用的是Anton de Bary(1879)的陈述:“是植物形成细胞,而不是细胞形成植物”(Sitte, 1992翻译)。从那时起,大量证据积累起来支持植物的有机体概念(参见Kaplan和Hagemann, 1991;Cooke和Lu, 1992;以及Kaplan, 1992;以及其中引用的文献)。
有机体理论特别适用于植物,其细胞在细胞分裂过程中不像动物细胞那样分开,而是最初通过插入细胞板(第4章)进行分隔。植物细胞的分离很少是完全的。相邻的植物细胞通过称为胞间连丝(plasmodesmata)的细胞质丝保持相互连接,这些丝穿过细胞壁并将整个植物体联合成一个有机整体。因此,植物被描述为超细胞生物(supracellular organisms)(Lucas等,1993)。
在现代形式中,细胞理论简单地陈述为:(1)所有生物体由一个或多个细胞组成,(2)生物体的化学反应,包括其能量相关过程和生物合成过程,发生在细胞内,(3)细胞来自其他细胞,(4)细胞含有它们所属生物体的遗传信息,并且这些信息从母细胞传递给子细胞。细胞理论和有机体理论并不相互排斥。它们共同提供了细胞和有机体水平上结构和功能的有意义视角(Sitte, 1992)。
“细胞(cell)”一词意为“小房间”,由Robert Hooke在17世纪引入,用于描述软木组织中由细胞壁分隔的小腔室。后来,Hooke认识到其他植物组织中的活细胞充满了“汁液”。最终,细胞的内容物被解释为生命物质,并被称为原生质(protoplasm)。认识到原生质复杂性的重要一步是Robert Brown在1831年发现了细胞核(nucleus)。这一发现很快伴随着细胞分裂(cell division)的报道。1846年,Hugo von Mohl注意到了原生质物质与细胞液(cell sap)之间的区别,1862年,Albert von Kölliker使用“细胞质(cytoplasm)”一词来指代细胞核周围的物质。细胞质中最显眼的包含物——质体,长期以来被认为仅仅是原生质的凝聚物。这些细胞器的独立身份和连续性的概念在19世纪确立。1880年,Johannes Hanstein引入了“原生质体(protoplast)”一词,用于指代细胞壁内的原生质单位。
每个活细胞都有一种将其内容物与外部环境隔离的手段。一种称为质膜(plasma membrane)或原生质膜(plasmalemma)的膜实现了这种隔离。此外,植物细胞在质膜外沉积了一层或多或少的刚性纤维素细胞壁(第4章)。质膜控制物质进出原生质体的通道,从而使细胞在结构和生化上与其周围环境有所不同。细胞内的过程可以释放和转移生长和维持代谢过程所需的能量。细胞被组织起来以保留和传递信息,使其自身及其后代的发展能够有序进行。这样,细胞所属的生物体的完整性得以维持。
在Hooke首次通过他的简易显微镜观察到软木结构的三个世纪以来,我们观察细胞及其内容物的能力有了显著的提高。随着光学显微镜的改进,现在可以观察到直径0.2微米(约200纳米)的物体,这比肉眼的分辨率提高了约500倍。透射电子显微镜(TEM)大大降低了光的分辨率限制。然而,由于标本制备、对比度和辐射损伤的问题,生物物体的分辨率更像是2纳米。尽管如此,这仍然比光学显微镜的分辨率好100倍。然而,TEM有明显的缺点:要观察的标本必须被保存(死亡)并切成非常薄的、实际上是二维的切片。使用荧光染料和各种照明方法的光学显微镜使生物学家能够克服这些问题,并在活植物细胞中观察亚细胞成分(Fricker和Oparka,1999;Cutler和Ehrhardt,2000)。值得注意的是使用来自水母Aequorea victoria的绿色荧光蛋白(GFP)作为荧光蛋白标签,以及使用共聚焦显微镜在完整组织中可视化荧光探针(Hepler和Gunning,1998;Fricker和Oparka,1999;Hawes等,2001)。在活植物细胞中观察亚细胞成分正在提供新的、常常是意想不到的关于亚细胞组织和动态的见解。
❙ 原核细胞和真核细胞
根据细胞内部组织的程度,现在识别出两种根本不同的生物群体:原核生物和真核生物。原核生物(pro,之前;karyon,核)由古菌和细菌代表,包括蓝细菌,而真核生物(eu,真;karyon,核)由所有其他生物代表(Madigan等,2003)。
原核细胞与真核细胞在遗传物质的组织上最为显著地不同。在原核细胞中,遗传物质是以一个大的、环状的脱氧核糖核酸(DNA)分子形式存在,与多种蛋白质松散结合。这个分子被称为细菌染色体,位于细胞质的一个区域,称为拟核(图2.1)。在真核细胞中,核DNA是线性的,并与称为组蛋白的特殊蛋白质紧密结合,形成一些更复杂的染色体。这些染色体被一个由两层膜组成的核膜包围,将它们与其他细胞内容物分开,形成一个明显的细胞核(图2.2)。原核细胞和真核细胞都含有蛋白质和核糖核酸(RNA)的复合物,称为核糖体,它们在从氨基酸亚基组装蛋白质分子的过程中起着关键作用。
图2.1
革兰氏阴性细菌Azotobacter vinelandii的电子显微照片。细胞质的颗粒状外观主要是由于存在大量的核糖体。较清晰的含DNA区域构成拟核。(由Jack L. Pate提供。)
真核细胞(eukaryotic cells)被膜(membranes)分隔成不同的区室(compartments),这些区室执行不同的功能。相比之下,原核细胞(prokaryotic cells)的细胞质(cytoplasm)通常不被膜分隔。值得注意的例外是蓝细菌(cyanobacteria)中广泛的光合膜系统(photosynthetic membranes)(类囊体(thylakoids))(Madigan等,2003),以及在多种细菌中发现的称为酸钙体(acidocalcisomes)的膜结合结构,包括引起冠瘿病(crown gall)的植物病原体——根癌农杆菌(Agrobacterium tumefaciens)(Seufferheld等,2003)。
在不同生物中,膜在电子显微镜下的外观非常相似。当适当保存和染色时,这些膜呈现三层结构,由两层暗层和中间一层较亮的层组成(图2.3)。这种类型的膜被Robertson(1962)命名为单位膜(unit membrane),并解释为两侧覆盖蛋白质层的双分子脂质层(bimolecular lipid layer)。尽管这种膜结构模型已被流动镶嵌模型(fluid mosaic model)所取代(见下文),但单位膜这一术语仍然是视觉上可定义的三层膜的有用名称。
真核细胞的内部膜包括围绕细胞核(nucleus)、线粒体(mitochondria)和质体的膜,这些是植物细胞的特征组成部分。真核细胞的细胞质还包含膜系统(内质网(endoplasmic reticulum)和高尔基体(Golgi apparatus))以及称为细胞骨架(cytoskeleton)的非膜性蛋白质丝(肌动蛋白丝(actin filaments)和微管(microtubules))的复杂网络。原核细胞中没有细胞骨架。植物细胞还发育出多功能细胞器,称为液泡(vacuoles),它们由称为液泡膜(tonoplast)的膜包围(图2.2)。
除了控制物质进出原生质体(protoplast)的质膜(plasma membrane)外,内部膜还控制物质在细胞内各区室之间的传递。通过这种方式,细胞可以维持不同细胞质区室中发生过程所需的特殊化学环境。膜还允许在细胞与其环境之间以及细胞相邻区室之间建立电位差或电压差。各种离子和分子的化学浓度差异以及跨膜电位差提供了用于驱动许多细胞过程的势能(potential energy)。
细胞内容物的区室化意味着在亚细胞水平上的分工(division of labor)。在多细胞生物中,分工也发生在细胞水平上,因为细胞分化并在某些功能上或多或少地特化。功能特化在细胞之间的形态差异中表现出来,这是多细胞生物结构复杂性的一个特征。
❙ 细胞质(CYTOPLASM)
如前所述,细胞质(cytoplasm)这一术语被引入,用以指代细胞核周围的原生质(protoplasmic)物质。随着时间的推移,人们在这种物质中发现了离散的实体,最初只是那些在光学显微镜分辨率范围内的实体;后来,随着电子显微镜的使用,发现了更小的实体。因此,细胞质的概念经历了演变;随着新技术的出现,这一概念无疑将继续发展。今天,大多数生物学家使用细胞质这一术语,正如Kölliker (1862)最初引入的那样,指代细胞核周围的所有物质,并将细胞质基质(cytoplasmic matrix)称为细胞溶质(cytosol),其中悬浮着细胞核、细胞器、膜系统和非膜实体。然而,最初定义的细胞溶质(cytosol)特指肝细胞中“除去线粒体和内质网成分的细胞质”(Lardy, 1965)。细胞质基质(cytoplasmic ground substance)和透明质(hyaloplasm)是植物细胞学家常用的术语,用以指代细胞质基质。一些生物学家将细胞质(cytoplasm)理解为细胞溶质(cytosol)。
在活的植物细胞中,细胞质始终处于运动状态;悬浮在细胞溶质中的细胞器和其他实体可以被观察到沿着移动的流有序地被带动。这种运动被称为细胞质流动(cytoplasmic streaming)或环流(cyclosis),是由肌动蛋白丝束(actin filaments)与所谓的运动蛋白(motor protein)——肌球蛋白(myosin)之间的相互作用引起的。肌球蛋白是一种含有ATP酶“头部”的蛋白质分子,其活性由肌动蛋白激活(Baskin, 2000; Reichelt and Kendrich-Jones, 2000)。细胞质流动是一个耗能的过程,无疑促进了细胞内(Reuzeau et al., 1997; Kost and Chua, 2002)以及细胞与其环境之间的物质交换。
图2.2
- Nicotiana tabacum(烟草)根尖。年轻细胞的纵切面。细节:er,内质网;m,线粒体;n,细胞核;ne,核膜;nu,核仁;o,油体;p,质体;v,液泡;w,细胞壁。(引自Esau, 1977。)
图2.3
电子显微照片显示了Allium cepa叶片两个细胞之间共同壁两侧的质膜(pm)的三层结构。壁两侧的微管(mt)在横切面视图中可见。
表2.1 ■ 植物细胞组分的清单
原生质体(protoplast)的各个组成部分将在以下段落中单独讨论。在这些组成部分中,有一种称为细胞器(organelle)的实体。与细胞质(cytoplasm)这个术语一样,细胞器这个术语也被不同的生物学家以不同的方式使用。一些人将细胞器这个术语的使用限制在膜结合实体上,如质体(plastid)和线粒体(mitochondria),而另一些人则更广泛地使用这个术语,也指内质网(endoplasmic reticulum)和高尔基体(Golgi body)以及非膜性成分,如微管(microtubule)和核糖体(ribosome)。本书中细胞器这个术语采用狭义的定义(表2.1)。
本章仅讨论质膜(plasma membrane)、细胞核(nucleus)和细胞质细胞器(cytoplasmic organelle)。原生质体的其余组成部分将在第3章中介绍。
❙ 质膜(PLASMA MEMBRANE)
在细胞的各种膜中,质膜在电子显微照片中通常具有最清晰的暗-亮-暗或单位膜(unit membrane)外观(图2.3; Leonard and Hodges, 1980; Robinson, 1985)。质膜有几个重要功能:(1)它介导物质进出原生质体的运输,(2)它协调细胞壁微纤维(纤维素)的合成和组装,(3)它转导参与控制细胞生长和分化的激素和环境信号。
质膜与细胞的内部膜具有相同的基本结构,由脂质双层(lipid bilayer)组成,其中嵌有球状蛋白质(globular protein),许多蛋白质横跨双层并在两侧突出(图2.4)。这些跨膜蛋白(transmembrane protein)嵌入双层中的部分是疏水的,而暴露在膜两侧的部分是亲水的。
图2.4
膜结构的流体镶嵌模型(fluid-mosaic model)。膜由脂质分子双层组成——它们的疏水"尾部"朝内——和大蛋白质分子。一些蛋白质(跨膜蛋白)横跨双层;其他蛋白质(外周蛋白,peripheral protein)附着在跨膜蛋白上。短碳水化合物链附着在质膜外表面大多数突出的跨膜蛋白上。整个结构相当流动;一些跨膜蛋白在双层内自由漂浮,并与脂质分子一起在其中横向移动,形成不同的图案或"镶嵌",因此蛋白质可以被认为漂浮在脂质"海洋"中。(来自Raven et al., 1992。)
膜的内外表面在化学组成上有显著差异。例如,植物细胞质膜中有两种主要的脂质——磷脂(含量较多)和甾醇(特别是stigmasterol)——而双层膜的两层中这些脂质的组成不同。此外,跨膜蛋白在双层膜中有明确的取向,突出在两侧的部分具有不同的氨基酸组成和三级结构。其他蛋白质也与膜相关,包括外周蛋白(peripheral proteins),之所以这样称呼是因为它们缺乏离散的疏水序列,因此不渗透到脂质双层中。跨膜蛋白和其他与膜紧密结合的脂质结合蛋白被称为整合蛋白(integral proteins)。在质膜的外表面,短链碳水化合物(寡糖,oligosaccharides)附着在突出的蛋白质上,形成糖蛋白(glycoproteins)。这些碳水化合物在某些真核细胞膜的外表面形成一层外衣,被认为在细胞间粘附过程和与细胞相互作用的分子(如激素、病毒和抗生素)的“识别”中起重要作用。
虽然脂质双层提供了细胞膜的基本结构和不透性,但蛋白质负责大多数膜功能。大多数膜由40%到50%的脂质(按重量计)和60%到50%的蛋白质组成,但膜中蛋白质的数量和类型反映了其功能。涉及能量转换的膜,如线粒体和叶绿体的内膜,由约75%的蛋白质组成。一些蛋白质是催化膜相关反应的酶,而另一些是参与特定分子进出细胞或细胞器的转运蛋白。还有一些作为受体,接收和转导来自细胞内部或外部环境的化学信号。虽然一些整合蛋白似乎被固定在某个位置(可能是细胞骨架),但脂质双层通常是相当流动的。一些蛋白质在双层中或多或少自由漂浮,它们和脂质分子可以在其中横向移动,形成不同的模式或镶嵌图案,这些图案随时间和地点而变化——因此这种膜结构模型被称为流动镶嵌模型(fluid-mosaic model)(图2.4;Singer和Nicolson,1972;Jacobson等,1995)。
膜含有不同类型的转运蛋白(Logan等,1997;Chrispeels等,1999;Kjellbom等,1999;Delrot等,2001)。其中两种类型是载体蛋白(carrier proteins)和通道蛋白(channel proteins),它们都只允许物质沿着其电化学梯度跨膜移动;也就是说,它们是被动转运体(passive transporters)。载体蛋白结合特定的被转运溶质,并经历一系列构象变化以将溶质转运过膜。通道蛋白形成水填充的孔,这些孔延伸穿过膜,当开放时,允许特定的溶质(通常是无机离子,如 K+、 Na+、Ca2+、Cl−)通过。通道不是持续开放的;相反,它们有“门”,这些门短暂打开然后再次关闭,这一过程被称为门控(gating)。
质膜(plasma membrane)和液泡膜(tonoplast)也含有称为水通道蛋白(aquaporins)的水通道蛋白,它们专门促进水通过膜的通道(Schäffner, 1998; Chrispeels et al., 1999; Maeshima, 2001; Javot and Maurel, 2002)。水相对自由地通过生物膜的脂双层(lipid bilayer),但水通道蛋白允许水更快地扩散通过质膜和液泡膜。由于液泡和细胞质(cytosol)必须保持恒定的渗透平衡(osmotic equilibrium),水的快速运动是必不可少的。有人认为,在高蒸腾作用(transpiration)期间,水通道蛋白促进了水从土壤快速流入根细胞并进入木质部。水通道蛋白已被证明可以阻止水在洪水期间流入根细胞(Tournaire-Roux et al., 2003),并在水稻的干旱回避(drought avoidance)中发挥作用(Lian et al., 2004)。此外,有证据表明,水通过水通道蛋白的运动增加是对某些诱导细胞扩张和生长的环境刺激的响应;质膜水通道蛋白的周期性表达与烟草的叶片展开机制有关(Siefritz et al., 2004)。
载体(carriers)可以根据其功能分为单向转运体(uniporters)和共转运体(cotransporters)。单向转运体仅将一种溶质(solute)从膜的一侧转运到另一侧。对于共转运体,一种溶质的转移取决于第二种溶质的同时或顺序转移。第二种溶质可以以相同方向转运,在这种情况下,载体蛋白被称为同向转运体(symporter),或者以相反方向转运,如反向转运体(antiporter)的情况。
物质逆其电化学梯度(electrochemical gradient)的转运需要能量输入,称为主动转运(active transport)。在植物中,这种能量主要由ATP驱动的质子泵(proton pump)提供,具体来说,是一种膜结合的H+-ATP酶(Sze et al., 1999; Palmgren, 2001)。该酶在膜上产生大量的质子梯度(proton gradient)(Pi+离子)。这种梯度为所有质子耦合共转运系统(proton-coupled cotransport systems)的溶质摄取提供了驱动力。液泡膜在植物膜中是独特的,因为它具有两种质子泵,一种H+-ATP酶和一种H+-焦磷酸酶(\nabla\primeH+-PPase)(Maeshima, 2001),尽管一些数据表明H+-PPase也可能存在于某些组织的质膜中(Ratajczak et al., 1999; Maeshima, 2001)。
大多数蛋白质和多糖(polysaccharides)等大分子的转运不能由转运离子和小极性分子(polar molecules)通过质膜的转运蛋白(transport proteins)来容纳。这些大分子通过从质膜出芽或与质膜融合的囊泡(vesicles)或囊状结构(saclike structures)进行转运,这一过程称为囊泡介导的转运(vesicle-mediated transport)(Battey et al., 1999)。通过从质膜出芽的囊泡将物质转运到细胞内的过程称为内吞作用(endocytosis),并涉及质膜的部分,称为被膜小窝(coated pits)(图2.5; Robinson
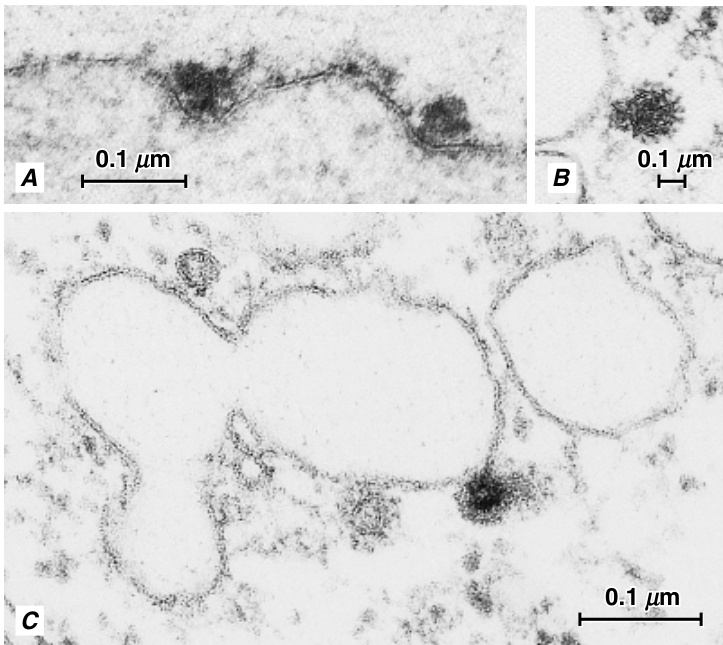{kind=link}
图2.5
暴露于含硝酸铅溶液中的玉米(Zea mays)根冠细胞的内吞作用(endocytosis)。A,在两个有被小窝(coated pits)中可以看到含铅的颗粒沉积物。B,一个含铅沉积物的有被小泡(coated vesicle)。C,这里,两个有被小泡中的一个已经与一个大的高尔基体小泡融合,并将释放其内容物。这个有被小泡(深色结构)仍然含有铅沉积物,但似乎已经失去了它的被膜,被膜位于其右侧。其左侧的有被小泡显然是完整的。(由David G. Robinson提供。)
以及Depta, 1988; Gaidarov et al., 1999。有被小窝是质膜上的凹陷,含有特定的受体(待运输到细胞内的分子必须先与其结合),并在其细胞质表面覆盖有网格蛋白(clathrin),这是一种由三条大肽链和三条小肽链组成的蛋白质,它们共同形成一个三叉结构,称为三脚蛋白复合体(triskelion)。有被小窝的内陷会掐断形成有被小泡。在细胞内,有被小泡会脱去其被膜,然后与其他膜结合结构(如高尔基体或小液泡)融合。通过小泡向相反方向的运输称为外排作用(exocytosis)(Battey et al., 1999)。在外排作用过程中,源自细胞内的小泡与质膜融合,将其内容物排出到细胞外。
在电镜制备的组织中,经常会遇到质膜的较大内陷或折叠。一些在细胞壁和原生质体之间形成囊袋,可能包括小管和小泡。一些内陷可能会推动液泡膜向前并侵入液泡。其他称为多泡体(multivesicular bodies)的结构,通常与质膜分离并嵌入细胞质中,或悬浮在液泡中。类似的构造最初在真菌中观察到,并命名为质膜外体(lomasomes)(Clowes和Juniper, 1968)。在烟草(Nicotiana tabacum) BY-2细胞中,多泡体已被确定为位于内吞途径上的植物前液泡区室,通向裂解液泡(见下文;Tse et al., 2004)。
❙ 细胞核(NUCLEUS)
通常是真核细胞原生质体中最显著的结构,细胞核执行两个重要功能:(1) 它通过决定细胞产生哪些RNA和蛋白质分子以及何时产生来控制细胞的持续活动,以及(2) 它是细胞大部分遗传信息的储存库,在细胞分裂过程中将其传递给子细胞。储存在细胞核中的总遗传信息称为核基因组(nuclear genome)。
细胞核由一对称为核膜(nuclear envelope)的膜所包围,两者之间存在核周间隙(perinuclear space)(图2.2和2.6; Dingwall和Laskey, 1992; Gerace和Foisner, 1994; Gant和Wilson, 1997; Rose等, 2004)。在许多地方,核膜的外膜与内质网(endoplasmic reticulum)相连,因此核周间隙与内质网的腔(lumen)是连续的。核膜被认为是内质网的一个特化的、局部分化的部分。核膜最显著的特征是存在大量圆柱形的核孔(nuclear pores),它们提供了细胞质(cytosol)与核基质(nucleoplasm)之间的直接接触(图2.6)。每个核孔周围的内膜和外膜相连,形成其开口的边缘。结构复杂的核孔复合体(nuclear pore complexes)——真核细胞中组装的最大的超分子复合体——跨越核孔处的核膜(Heese-Peck和Raikhel, 1998; Talcott和Moore, 1999; Lee, J.-Y.等, 2000)。核孔复合体大致呈轮状,部分由一个圆柱形的中央通道(轮毂)组成,从该通道向外伸出八个辐条,与核孔内衬的核膜相连的互锁环相连。核孔复合体允许某些离子和小分子通过扩散通道相对自由地通过,这些通道的直径约为9纳米。通过核孔复合体运输的蛋白质和其他大分子大大超过了这个通道尺寸。它们的运输是通过一个高度选择性的主动(能量依赖)运输机制进行的,该机制通过中央通道进行。中央通道的功能直径可达26纳米(Hicks和Raikhel, 1995; Görlich和Mattaj, 1996; Görlich, 1997)。
图2.6
核膜(ne)的剖面图\eta(A)和表面图(B,中央部分)显示核孔(po)。A图中核孔中的电子致密物质在B图中显示为具有中央颗粒的环状结构。A图中膜之间的透明空间称为核周间隙。来自Mimosa pudica叶柄的薄壁细胞。(来自Esau, 1977。)
在特殊染色的细胞中,染色质(chromatin)的细丝和颗粒可以从核质(nucleoplasm)中区分出来。染色质由DNA与大量称为组蛋白(histones)的蛋白质结合而成。在核分裂过程中,染色质逐渐变得更加浓缩,直到形成染色体(chromosomes)。不分裂或间期(interphase)的细胞核中的染色体(染色质)在一个或多个位点附着在核膜(nuclear envelope)的内膜上。在DNA复制之前,每条染色体都由一条长的DNA分子组成,携带遗传信息。在大多数间期细胞核中,大部分染色质是弥散的,染色较浅。这种未浓缩的染色质称为常染色质(euchromatin),具有遗传活性,与高水平的RNA合成相关。剩余的浓缩染色质称为异染色质(heterochromatin),不具有遗传活性;也就是说,它与RNA合成无关(FrankliNand Cande, 1999)。总体而言,只有一小部分染色体DNA编码必需的蛋白质或RNA;显然,高等生物的基因组中存在大量的DNA冗余(Price, 1988)。细胞核中可能含有未知功能的蛋白质内含物,呈结晶状、纤维状或无定形(Wergin et al., 1970),此外还有含有染色质的“微泡”(micropuffs)和由核糖核蛋白(ribonucleoprotein)组成的卷曲体(coiled bodies)(Martín et al., 1992)。
不同生物的体细胞(somatic cells)(营养细胞或体细胞)中的染色体数量各不相同。Haplopappus gracilis,一种沙漠一年生植物,每个细胞有4条染色体;Arabidopsis thaliana,10条;Vicia faba,蚕豆,12条;Brassica oleracea,卷心菜,18条;Asparagus officinalis,20条;Triticum vulgare,面包小麦,42条;Cucurbita maxima,南瓜,48条。生殖细胞(gametes)或配子(gametes)的染色体数量仅为生物体体细胞特征数量的一半。配子中的染色体数量称为单倍体(haploid)(单套)数量,用n表示,而体细胞中的染色体数量称为二倍体(diploid)(双套)数量,用2n表示。具有超过两套染色体的细胞称为多倍体(polyploid)(3n、4n、5n或更多)。
通常,在光学显微镜下,细胞核内唯一可见的结构是称为核仁(nucleoli)(单数:nucleolus)的球形结构(图2.2;Scheer et al., 1993)。核仁含有高浓度的RNA和蛋白质,以及从几条染色体延伸出的大环DNA。这些DNA环称为核仁组织区(nucleolar organizer regions),含有核糖体RNA(rRNA)基因簇。在这些位点,新形成的rRNA与从细胞质(cytosol)输入的核糖体蛋白一起包装,形成核糖体亚基(大亚基和小亚基)。然后,核糖体亚基通过核孔(nuclear pores)转移到细胞质中,在那里组装成核糖体。尽管核仁通常被认为是核糖体制造的场所,但它只参与了这一过程的一部分。核仁的存在是由于正在包装以形成核糖体亚基的分子积累所致。
在许多二倍体生物中,细胞核中每个单倍体染色体组含有一个核仁(nucleolus)。核仁可能会融合,然后表现为一个大的结构。核仁的大小反映了其活性水平。除了核仁组织区(nucleolar organizer region)的DNA外,核仁还含有由已经与蛋白质结合形成纤维的rRNA组成的纤维成分(fibrillar component),以及由成熟的核糖体亚基组成的颗粒成分(granular component)。活跃的核仁还显示出通常被称为液泡(vacuoles)的浅染色区域。在活体培养细胞中,这些区域(不应与细胞质中膜结合的液泡混淆)可以看到反复收缩的现象,这可能与RNA运输有关。
核分裂(nuclear division)有两种类型:有丝分裂(mitosis),在此期间,一个细胞核产生两个子细胞核,每个子细胞核在形态和遗传上都与另一个子细胞核和亲本细胞核相同;减数分裂(meiosis),在此期间,亲本细胞核经历两次分裂,其中一次是减数分裂。通过精确的机制,减数分裂产生四个子细胞核,每个子细胞核的染色体数量是亲本细胞核的一半。在植物中,有丝分裂产生体细胞(somatic cells)和配子(gametes)(精子和卵子),而减数分裂产生减数孢子(meiospores)。在这两种分裂中(有一些例外),核膜(nuclear envelope)会破裂成碎片,这些碎片变得与内质网(ER)池无法区分,核孔复合体(nuclear pore complexes)也会被分解。当在末期(telophase)组装新的细胞核时,内质网囊泡(ER vesicles)会结合形成两个核膜,并形成新的核孔复合体(Gerace和Foisner,1994)。核仁在前期(prophase)晚期(有一些例外)分散,并在末期重新组织。
❙ 细胞周期(CELL CYCLE)
活跃分裂的体细胞通过一系列称为细胞周期(cell cycle)的常规事件。细胞周期通常分为间期(interphase)和有丝分裂(mitosis)(图2.7;Strange,1992)。间期在有丝分裂之前,在大多数细胞中,有丝分裂之后是细胞质分裂(cytokinesis),即细胞质部分的分裂和子细胞核分离成单独的细胞(第4章)。因此,大多数植物细胞是单核的。某些特化细胞可能只在发育过程中(例如,核型胚乳(nuclear endosperm))或终生(例如,非节状乳汁管(nonarticulated laticifers))成为多核的。有丝分裂和细胞质分裂一起被称为细胞周期的M期(M phase)。
图2.7
细胞周期。细胞分裂由有丝分裂(细胞核的分裂)和细胞质分裂(细胞质的分裂)组成,发生在间期的三个准备阶段(G1、\DeltaS和 G2)完成之后。细胞周期的进展主要在两个检查点(checkpoints)控制,一个在G1结束时,另一个在G2结束时。G2期之后是有丝分裂,通常随后是细胞质分裂。有丝分裂和细胞质分裂一起构成细胞周期的M期。在不同物种或同一生物体内不同组织的细胞中,各个阶段占整个周期的比例不同。(来自Raven等,2005。)
间期(interphase)可以分为三个阶段,分别称为G1期、S期和G2期。G1期(G代表间隙(gap))发生在有丝分裂(mitosis)之后。这是一个生化活动旺盛的时期,在此期间细胞体积增大,各种细胞器(organelle)、内膜(internal membrane)和其他细胞质(cytoplasmic)成分的数量增加。S期(合成(synthesis)期)是DNA复制的时期。在DNA复制开始时,二倍体(diploid)细胞核的DNA含量为2C(C是单倍体(haploid)DNA含量);在S期结束时,DNA含量翻倍至4C。在S期,许多组蛋白(histone)和其他与DNA相关的蛋白质也被合成。S期之后,细胞进入G2期,G2期在S期之后,在有丝分裂之前。S期的主要作用是确保染色体复制完成,并允许修复受损的DNA。前前期带(preprophase band)的微管(microtubule)也在G2期发育,前前期带是一个环状的微管带,位于质膜(plasma membrane)边缘,并在与细胞分裂平面相对应的平面上环绕细胞核(第4章;Gunning和Sammut,1990)。在有丝分裂期间,S期合成的遗传物质被平均分配到两个子细胞核中,恢复2C的DNA含量。
在所有真核细胞(eukaryotic cell)中,调控细胞周期的控制机制的性质基本上相似。在典型的细胞周期中,进程在关键的过渡点(称为检查点(checkpoint))受到控制——首先在G1-S期过渡点,然后在 G2-M过渡点(Boniotti和Griffith,2002)。第一个检查点决定细胞是否进入S期,第二个检查点决定是否启动有丝分裂。第三个检查点,即中期(metaphase)检查点,如果某些染色体没有正确附着在有丝分裂纺锤体(mitotic spindle)上,则会延迟后期(anaphase)。细胞周期的进程取决于检查点上细胞周期蛋白依赖性蛋白激酶(cyclin-dependent protein kinase, CDK)的成功形成、激活和随后的失活。这些激酶由催化CDK亚基和激活的细胞周期蛋白(cyclin)亚基组成(Hemerly等,1999;Huntley和Murray,1999;Mironov等,1999;Potuschak和Doerner,2001;Stals和Inzé,2001)。生长素(auxin)和细胞分裂素(cytokinin)都被认为参与了植物细胞周期的调控(Jacqmard等,1994;Ivanova和Rost,1998;den Boer和Murray,2000)。
处于G1期的细胞有几种选择。在足够的刺激下,它们可以继续进行细胞分裂并进入S期。它们可能会因环境因素(如冬季休眠)而暂停细胞周期的进程,并在稍后恢复分裂。这种特殊的静止或休眠状态通常称为Go期(G-zero期)。其他命运包括分化(differentiation)和程序性细胞死亡(programmed cell death),这是一种遗传决定的程序,可以协调细胞的死亡(第5章;Lam等,1999)。
某些细胞仅进行DNA复制和间隙期(gap phases),而不进行后续的核分裂,这一过程称为内复制(endoreduplication)(第5章;D’Amato, 1998;Larkins et al., 2001)。单个细胞核随后变为多倍体(内多倍体,endopolyploidy或endoploidy)。内多倍体可能是单个细胞分化的组成部分,例如在Arabidopsis的毛状体(trichome)中(第9章),也可能是任何组织或器官的分化组成部分。在大多数植物细胞中,细胞体积与多倍体程度之间存在正相关关系,表明多倍体核可能是形成大型植物细胞所必需的(Kondorosi et al., 2000)。
❙ 质体
与液泡(vacuoles)和细胞壁(cell walls)一起,质体是植物细胞的特征性组成部分(Bowsher and Tobin, 2001)。每个质体被由两层膜组成的包膜(envelope)包围。质体内部被分化为一个或多或少均质的基质(stroma)和一个称为类囊体(thylakoids)的膜系统。细胞质(cytosol)和质体基质之间的主要渗透屏障是质体包膜的内膜。外膜虽然对细胞质蛋白具有屏障作用,但通常被认为对小分子溶质(<600 Da)是可渗透的,这一假设可能需要重新评估(Bolter and Soll, 2001)。在一些质体表面观察到充满基质的管状结构。
这些所谓的基质管(stromules)可以连接不同的质体,并且已被证明允许绿色荧光蛋白(green fluorescent protein)在质体之间交换(Kohler et al., 1997;Köhler and Hanson, 2000;Arimura et al., 2001;Gray et al., 2001;Pyke and Howells, 2002;Kwok and Hanson, 2004)。在一项关于基质管生物发生的研究中,基质管长度和频率的增加与有色体(chromoplast)分化相关;研究提出,基质管增强了质体的特定代谢活性(Waters et al., 2004)。
质体是半自主的细胞器,广泛认为它们是通过内共生(endosymbiosis)过程从自由生活的蓝细菌(cyanobacteria)进化而来的(Palmer and Delwiche, 1998;Martin, 1999;McFadden, 1999;ReumanNand Keegstra, 1999;Stoebe and Maier, 2002)。事实上,质体在多个方面与细菌相似。例如,质体与细菌一样含有核样体(nucleoids),这些区域含有DNA。质体的DNA与细菌的DNA一样以环状形式存在(Sugiura, 1989);此外,它不与组蛋白(histones)结合。在进化过程中,内共生体(蓝细菌)的大部分DNA逐渐转移到宿主细胞核中;因此,现代质体的基因组与核基因组相比非常小(Bruce, 2000;RujaNand Martin, 2001)。质体和细菌都含有核糖体(70S核糖体),其大小约为细胞质中与内质网(endoplasmic reticulum)相关的核糖体(80S核糖体)的三分之二。(S代表Svedbergs,即沉降系数的单位。)此外,质体分裂的过程——二分裂(binary fission)——在形态上与细菌细胞分裂相似。
叶绿体含有叶绿素和类胡萝卜素色素
成熟的质体通常根据它们所含的色素种类进行分类。叶绿体(Chloroplasts)(图2.8-2.10)是光合作用的场所,含有叶绿素和类胡萝卜素色素。叶绿素色素使这些质体呈现绿色,它们存在于绿色植物部位,在叶片中数量特别多且分化良好。在种子植物中,叶绿体通常呈盘状,直径在4到6微米之间。单个叶肉(mesophyll)细胞中叶绿体的数量差异很大,取决于物种和细胞的大小(Gray, 1996)。可可(Cacao theobroma)和Peperomia metallia叶片的单个叶肉细胞可能只含有3个叶绿体,而萝卜(Raphanus sativus)叶片的单个叶肉细胞中可能含有多达300个叶绿体。大多数被研究过质体发育的叶片,其叶肉细胞每个含有50到150个叶绿体。叶绿体通常
图2.8
叶绿体的三维结构。注意内部膜(类囊体)不与质体被膜相连。(来自Raven等, 1992。)
发现它们的宽面与细胞壁平行,优先位于与气室相邻的细胞表面。它们可以在光的影响下在细胞内重新定向——例如,在低或中等光强下聚集在与叶表面平行的壁附近,从而优化光合作用的光利用(Trojan和Gabrys, 1996; Williams等, 2003)。在可能造成损害的高光强下,叶绿体可以沿着与叶表面垂直的壁定向。光谱的蓝-紫外区域是叶绿体运动最有效的刺激(Trojan和Gabrys, 1996; Yatsuhashi, 1996; Kagawa和Wada, 2000, 2002)。在黑暗中,叶绿体要么随机分布在所有细胞壁周围,要么它们的排列取决于细胞内的局部因素(Haupt和Scheuerlein, 1990)。推测叶绿体的运动涉及一个基于肌动蛋白-肌球蛋白的系统。
叶绿体的内部结构很复杂。基质被一个精细的类囊体系统穿过,该系统由基粒(grana)(单数:granum)——类似于一叠硬币的盘状类囊体堆叠——和基质类囊体(或基粒间类囊体)组成,后者穿过基粒之间的基质并将它们相互连接(图2.8-2.10)。基粒和基质类囊体及其内部隔室被认为构成一个单一的、相互连接的系统。类囊体与质体被膜没有物理连接,而是完全嵌入基质中。叶绿素和类胡萝卜素色素——两者都参与光捕获——与蛋白质一起嵌入类囊体膜中,形成称为光系统的离散组织单元。类胡萝卜素色素的主要功能是作为抗氧化剂,防止叶绿素分子的光氧化损伤(Cunningham和Gantt, 1998; Vishnevetsky等, 1999; Niyogi, 2000)。
叶绿体(chloroplast)通常含有淀粉(starch)、植物铁蛋白(phytoferritin,一种铁化合物)和以球状体形式存在的脂质(lipid),称为质体小球(plastoglobuli,单数:plastoglobule)。淀粉粒是暂时的储存产物,只有在植物进行光合作用时才会积累。在黑暗中仅24小时的植物叶绿体中可能缺乏淀粉粒,但在光照下仅3或4小时后,淀粉粒通常会重新出现。
成熟的叶绿体含有多个环状质体DNA分子的拷贝,以及复制、转录和翻译该遗传物质的机制(Gray, J. C., 1996)。然而,由于叶绿体的编码能力有限(约100种蛋白质),绝大多数与叶绿体生物发生和功能相关的蛋白质都是由核基因组编码的(Fulgosi and Soll, 2001)。这些蛋白质在细胞质中的核糖体上合成,作为前体蛋白质,借助称为转运肽(transit peptide)的氨基末端延伸被靶向到叶绿体中。每个导入叶绿体的蛋白质都含有特定的转运肽。转运肽不仅将蛋白质靶向到叶绿体,还介导其导入基质(stroma),在导入后转运肽被切除(Flügge, 1990; Smeekens et al., 1990; Theg and Scott, 1993)。跨类囊体膜(thylakoid membrane)的运输由第二个转运肽介导,当第一个转运肽被切除时,第二个转运肽被暴露(Cline et al., 1993; Keegstra and Cline, 1999)。有证据表明,部分叶绿体蛋白质机制来源于叶绿体的内共生蓝细菌祖先(ReumanNand Keegstra, 1999; Bruce, 2000)。
除了从细胞核到叶绿体的调节性运输外,叶绿体还向细胞核传递信号,以协调核和叶绿体基因的表达。此外,质体信号还调节非质体蛋白质的核基因表达以及线粒体基因的表达(见Rodermel, 2001中的参考文献)。叶绿体不仅是光合作用的场所,还参与氨基酸合成和脂肪酸合成,并为淀粉的暂时储存提供空间。
色质体仅含有类胡萝卜素色素
色质体(chromoplast,chroma意为颜色)也是有色质体(pigmented plastid)(图2.11)。它们的形状多变,缺乏叶绿素(chlorophyll),但合成并保留类胡萝卜素(carotenoid)色素,这些色素通常是许多花朵、老叶、一些果实和一些根部的黄色、橙色或红色的原因。色质体是最异质的质体类别,完全根据存在的类胡萝卜素成分的结构进行分类。
图2.9
A,荠菜(Capsella bursa-pastoris)叶细胞中沿细胞壁分布的叶绿体。线粒体(m)在空间上与叶绿体紧密相关。B,烟草(Nicotiana tabacum)叶片中可见的叶绿体及其基粒(grana)的剖面图。(B,来自Esau, 1977。)
在成熟的质体(plastid)中(Sitte et al., 1980)。大多数属于以下四种类型之一:(1)球状有色体(globular chromoplasts),具有许多含有类胡萝卜素的质体小球(plastoglobuli)(图2.11A)。类囊体(thylakoid)的残余也可能存在。质体小球通常集中在被膜(envelope)下的外周基质(stroma)中(如毛茛属(Ranunculus repens)的花瓣和辣椒属(Capsicum)的黄色果实、郁金香属(Tulipa)的花被、柑橘属(Citrus)的果实);(2)膜状有色体(membranous chromoplasts),其特征是具有多达20层同心(双层)含类胡萝卜素的膜(图2.11B)(如水仙属(Narcissus)和甜橙(Citrus sinensis)的花瓣);(3)管状有色体(tubular chromoplasts),其中类胡萝卜素被整合到丝状脂蛋白"小管"中(图2.11C)(如辣椒属(Capsicum)的红色果实、蔷薇属(Rosa)的花托、旱金莲属(Tropaeolum)的花瓣;Knoth et al., 1986);(4)结晶有色体(crystalline chromoplasts),其含有纯胡萝卜素的结晶内含物(图2.11D)(如胡萝卜(Daucus)根中的\Delta[3-胡萝卜素和番茄(Solanum lycopersicum)果实中的番茄红素)。胡萝卜素晶体通常被称为色素体(pigment bodies),起源于类囊体(thylakoids),并在发育的所有阶段都保持被质体被膜(plastid envelope)包围。
图2.10
叶绿体结构。A,在光学显微镜下,叶绿体中的基粒(grana)呈现为点状。这些叶绿体来自番茄(Solanum lycopersicum)的子叶。B,玉米(Zea)叶片维管束鞘细胞中叶绿体的电子显微照片,显示表面的基粒。(A,来自Hagemann, 1960。)
球状有色体是最常见的类型,在进化上被认为是最古老和最原始的(Camara et al., 1995)。
有色体可能通过转化从先前存在的绿色叶绿体发育而来,在此过程中叶绿体的叶绿素和类囊体膜消失,类胡萝卜素大量积累,如许多果实成熟时发生的情况(Ziegler et al., 1983; Kuntz et al., 1989; Marano and Carrillo, 1991, 1992; Cheung et al., 1993; Ljubesic et al., 1996)。有趣的是,这些变化显然伴随着质体核糖体(plastid ribosomes)和rRNA的消失,但质体DNA保持不变(Hansmann et al., 1987; Camara et al., 1989; Marano and Carrillo, 1991)。随着质体核糖体和rRNA的丢失,有色体中不能再进行蛋白质合成,这表明有色体特异性蛋白质必须在细胞核中编码,然后导入到发育中的有色体中。然而,有色体的发育并不是不可逆的现象。例如,柑橘类果实(Goldschmidt, 1988)和胡萝卜根(Grönegress, 1971)的有色体能够逆向分化为叶绿体;它们失去胡萝卜素色素,并发育出类囊体系统、叶绿素和光合作用装置。
有色体的确切功能尚不清楚,尽管有时它们作为吸引昆虫和其他与其共同进化的动物的引诱剂,在开花植物的异花授粉和果实及种子的传播中发挥重要作用(Raven et al., 2005)。
白色体(Leucoplasts)是无色质体
在结构上,白色体(leucoplasts)(图2.12)是成熟质体中最不分化的,通常具有均匀的颗粒状基质(stroma)、多个类核(nucleoids),尽管有相反的报道,但它们具有典型的70S核糖体(70S ribosomes)。它们缺乏复杂的内膜系统(inner membranes)(Carde, 1984; Miernyk, 1989)。一些白色体储存淀粉(淀粉体(amyloplasts);图2.13),另一些储存蛋白质(蛋白质体(proteinoplasts))、脂肪(油体(elaioplasts))或这些物质的组合。淀粉体被分类为简单或复合淀粉体(Shannon, 1989)。简单淀粉体,如马铃薯块茎中的淀粉体,含有单个淀粉粒,而复合淀粉体则含有多个通常紧密堆积的淀粉粒,如燕麦和水稻的胚乳中。马铃薯块茎中的淀粉粒可能变得非常大,以至于包膜破裂(Kirk和Tilney-Bassett, 1978)。根冠中的复合淀粉体在重力感知中起着至关重要的作用(Sack和Kiss, 1989; Sack, 1997)。
所有质体最初都来源于前质体
前质体(proplastids)是小的、无色的质体,存在于植物体的未分化区域,如根和茎的顶端分生组织(apical meristems)(Mullet, 1988)。合子(zygotes)含有前质体,这些前质体是成年植物中所有质体的最终前体。在大多数被子植物中,合子的前质体完全来自卵细胞的细胞质(Nakamura等, 1992)。然而,在针叶树中,合子的前质体来自精子细胞携带的前质体。无论哪种情况,结果都是个体植物的质体基因组通常是从单一亲本遗传的。由于成年植物中的所有质体都来自单一亲本,因此个体植物内的所有质体(无论是叶绿体(chloroplasts)、有色体(chromoplasts)还是白色体)都具有相同的基因组(dePamphilis和Palmer, 1989)。每个前质体含有一个单一的环状DNA分子。
图2.12
Zebrina叶表皮细胞中聚集在细胞核周围的白色体。(×620.)
图2.13
大豆(Glycine max)胚囊中的淀粉体,一种白色体。圆形、透明的体是淀粉粒。较小的、致密的体是油体。淀粉体参与种子和储存器官(如马铃薯块茎)中淀粉的合成和长期储存。(由Roland R. Dute提供。)
如前所述,质体(plastid)通过二分裂(binary fission)进行繁殖,这是一种将细胞分成两半的过程,是细菌的特征(Oross and Possingham, 1989)。在分生组织细胞(meristematic cells)中,前质体(proplastid)的分裂大致与细胞分裂保持同步。
图2.14
- Beta vulgaris叶片中正在分裂的叶绿体。如果分裂过程继续下去,两个子代质体将在狭窄的收缩处或峡部(isthmus)分离。在收缩处的右侧可以看到三个过氧化物酶体(peroxisome)。
前质体必须在细胞分裂之前进行分裂。成熟细胞中的质体数量通常超过原始前质体的数量。最终质体数量的较大部分可能来自细胞扩展期间成熟质体的分裂。尽管质体分裂显然是由细胞核控制的(Possingham and Lawrence, 1983),但质体DNA复制和质体分裂之间存在密切的相互作用。
质体分裂是由质体中间的收缩引发的(图2.14)。随着收缩的持续变窄,两个子代质体通过一个狭窄的峡部连接,最终断裂。然后,子代质体的被膜(envelope membranes)重新密封。收缩过程是由称为质体分裂环(plastid-dividing rings)的收缩环引起的,这些环在电子显微镜下可见为电子致密带。有两个同心的质体分裂环,一个外环位于质体外膜的胞质面(cytosolic face),一个内环位于质体内膜的基质面(stromal face)。
图2.15
甘蔗(Saccharum officinarum)叶细胞中具有前片层体(prolamellar body)的黄化叶绿体。质体中的核糖体(ribosome)很明显。(由W. M. Laetsch提供)
图2.16
在质体分裂环出现之前,两种类似细胞骨架的蛋白质(citoskeletal-like proteins),FtsZ1和FtsZ2——细菌细胞分裂FtsZ蛋白的同源物——在质体被膜内的基质(stroma)中未来的分裂位点组装成一个环。有人认为FtsZ环决定了分裂区域(Kuroiwa et al., 2002)。叶绿体分裂的分子分析表明,质体分裂的机制是从细菌细胞分裂进化而来的(Osteryoung and Pyke, 1998; Osteryoung and McAndrew, 2001; Miyagishima et al., 2001)。
如果前质体(proplastid)向更高分化形式发育的过程因缺乏光照而受阻,它可能会形成一个或多个原片层体(prolamellar body)(图2.15),原片层体是由管状膜组成的准晶体结构(Gunning, 2001)。含有原片层体的质体称为黄化质体(etioplast)(Kirk和Tilney-Bassett, 1978)。黄化质体在黑暗条件下生长的植物叶片细胞中形成。在随后黄化质体在光照下发育成叶绿体的过程中,原片层体的膜发育成类囊体(thylakoid)。已证明类胡萝卜素(carotenoid)的合成是拟南芥(Arabidopsis)黄化幼苗中原片层体形成所必需的(Park等, 2002)。在自然界中,一些种子胚胎中的前质体首先发育成黄化质体;然后在光照下,黄化质体发育成叶绿体。各种类型的质体因其相对容易从一种类型转变为另一种类型而引人注目(图2.16)。
质体发育周期,从叶绿体由前质体发育开始(A)。最初,前质体含有很少或没有内膜。B-D,随着前质体分化,扁平囊泡从质体被膜的内膜发育而来,最终排列成基粒(grana)和基质类囊体(stroma thylakoid)。E,成熟叶绿体的类囊体系统与被膜不连续。F,G,前质体也可能发育成有色体(chromoplast)和白色体(leucoplast)。这里展示的白色体是合成淀粉的造粉体(amyloplast)。请注意,有色体可以由前质体、叶绿体或白色体形成。各种类型的质体可以从一种类型转变为另一种类型(虚线箭头)。(摘自Raven等, 2005)
❙ 线粒体
线粒体(mitochondrion)与质体一样,由两层膜包围(图2.17和2.18)。内膜向内折叠形成许多称为嵴(crista)(单数:crista)的褶皱,这大大增加了酶及其相关反应可利用的表面积。线粒体通常比质体小,直径约为半微米,长度和形状变化很大。
线粒体是呼吸作用(respiration)的场所,呼吸作用涉及从有机分子中释放能量并将其转化为ATP(三磷酸腺苷)分子。
图2.17
线粒体的三维结构。包围线粒体的两层膜中的内膜向内折叠,形成嵴。许多参与呼吸作用的酶和电子载体存在于嵴中。(摘自Raven等, 2005)
图2.18
线粒体。A,在烟草(Nicotiana tabacum)叶细胞中。被膜由两层膜组成,嵴嵌入致密的基质中。B,菠菜(Spinacia oleracea)叶细胞中的线粒体,切片显示类核(nucleoid)中的一些DNA链。细节:cw,细胞壁。
三磷酸腺苷(ATP),是细胞的主要即时能量来源(Mackenzie and McIntosh, 1999; Møller, 2001; Bowsher and Tobin, 2001)。在最内层的隔室中,围绕嵴的是基质,一种含有酶、辅酶、水、磷酸盐和其他与呼吸有关的分子的密集溶液。虽然外膜对大多数小分子相当通透,但内膜相对不透,只允许某些分子(如丙酮酸和ATP)通过,而阻止其他分子的通过。一些柠檬酸循环的酶存在于基质溶液中。其他柠檬酸循环酶和电子传递链的组分则嵌入嵴的表面。大多数植物细胞含有数百个线粒体,每个细胞中线粒体的数量与细胞对ATP的需求有关。
线粒体处于不断运动中,似乎在流动的细胞质中自由移动,从一个细胞部分移动到另一个部分;它们也通过二分裂(binary fission)融合和分裂(Arimura et al., 2004),涉及类似于质体分裂环(plastid-dividing rings)的分裂环(Osteryoung, 2000)。在烟草(Nicotiana tabacum)培养细胞中,线粒体的运动已被证明涉及基于肌动蛋白-肌球蛋白的系统(Van Gestel et al., 2002)。线粒体倾向于聚集在需要能量的地方。在细胞膜非常活跃地运输物质进出细胞的细胞中,线粒体通常可以沿着膜表面排列。
线粒体,像质体一样,是半自主的细胞器,含有合成一些自身蛋白质所需的组分。在基质中发现一个或多个含DNA的核样体和许多类似于细菌的70S核糖体(图2.18)。DNA不与组蛋白结合。因此,在植物细胞中,遗传信息存在于三个不同的隔室中:细胞核、质体和线粒体。植物的线粒体基因组(200-2400 kb)比动物(14-42 kb)、真菌(18-176 kb)和质体(120-200 kb)的基因组大得多(Backert et al., 1997; Giegé and Brennicke, 2001)。它们的结构组织尚未完全理解。不同大小的线性和环状DNA分子以及更复杂的DNA分子始终存在(Backert et al., 1997)。
线粒体被广泛认为是通过内共生(endosymbiosis)过程从自由生活的α-变形菌(α-proteobacteria)进化而来的(Gray, 1989)。与叶绿体一样,在进化过程中,线粒体的DNA大量转移到细胞核中(Adams et al., 2000; Gray, 2000)。证据还表明,一些遗传信息在长时间的进化过程中从叶绿体转移到线粒体(Nugent and Palmer, 1988; Jukes and Osawa, 1990; Nakazono and Hirai, 1993),并可能从细胞核转移到线粒体(Schuster and Brennicke, 1987; Marienfeld et al., 1999)。植物线粒体基因组仅编码约30种蛋白质。相比之下,估计约有4000种由细胞核编码的蛋白质从细胞质中导入。核编码的线粒体蛋白质在其N端含有称为前序列(presequences)的信号肽,以引导它们进入线粒体(BrauNand Schmitz, 1999; Mackenzie and McIntosh, 1999; Giegé and Brennicke, 2001)。
仅在线粒体DNA中发现的遗传信息可能对细胞发育有影响。最显著的是细胞质雄性不育(cytoplasmic male sterility),这是一种母系遗传(mitochondrial DNA is maternally inherited)的性状,它阻止功能性花粉的产生,但不影响雌性生育能力(Leaver and Gray, 1982)。由于它阻止自花授粉,细胞质雄性不育表型已被广泛用于F1杂交种子的商业化生产(例如,在玉米、洋葱、胡萝卜、甜菜和矮牵牛中)。
线粒体已被视为动物细胞中程序性细胞死亡(programmed cell death),即凋亡(apoptosis)调控的关键参与者(第5章; Desagher and Martinou, 2000; Ferri and Kroemer, 2001; Finkel, 2001)。凋亡的主要细胞触发因素是细胞色素c从线粒体膜间隙释放。细胞色素c的释放似乎是激活称为caspases(凋亡特异性半胱氨酸蛋白酶)的分解代谢蛋白酶的关键事件。尽管线粒体可能在植物程序性细胞死亡中起作用,但释放的细胞色素c不太可能参与该过程(Jones, 2000; Xu and Hanson, 2000; Young and Gallie, 2000; Yu et al., 2002; Balk et al., 2003; Yao et al., 2004)。
❙ 过氧化物酶体(peroxisomes)
与由两层膜包围的质体和线粒体不同,过氧化物酶体(peroxisomes)(也称为微体(microbodies))是由单层膜包围的球形细胞器(图2.14和2.19; Frederick et al., 1975; Olsen, 1998)。然而,过氧化物酶体与质体和线粒体最显著的区别在于它们缺乏DNA和核糖体。因此,所有过氧化物酶体蛋白都是由细胞核编码的,至少基质蛋白(matrix proteins)是在细胞质中的核糖体上合成,然后转运到过氧化物酶体中。一部分过氧化物酶体膜蛋白可能首先靶向内质网(endoplasmic reticulum),然后通过囊泡介导的运输(vesicle-mediated transport)从那里转运到细胞器(JohnsoNand Olsen, 2001)。过氧化物酶体的大小范围为0.5至1.5\upmum。它们缺乏内膜,内部呈颗粒状,有时含有由蛋白质组成的无定形或结晶体。根据主流观点,过氧化物酶体是自我复制的细胞器,新的过氧化物酶体通过分裂从已有的过氧化物酶体产生。从内质网到过氧化物酶体的囊泡介导途径的存在使一些研究人员推测,这些细胞器也可能从头生成(de novo)(Kunau and Erdmann, 1998; Titorenko and Rachubinski, 1998; Mullen et al., 2001),这一观点受到其他人的强烈质疑(Purdue and Lazarow, 2001)。在生化上,过氧化物酶体的特征是至少存在一种产生过氧化氢的氧化酶(oxidase)和用于去除过氧化氢的过氧化氢酶(catalase)(Tolbert, 1980; Olsen, 1998)。正如Corpas等人(2001)所指出的,过氧化物酶体的一个重要特性是它们的"代谢可塑性(metabolic plasticity)",即它们的酶含量可以变化,取决于生物体、细胞类型或组织类型以及环境条件。过氧化物酶体执行多种代谢功能(Hu et al., 2002)。
在植物中,两种非常不同类型的过氧化物酶体(peroxisome)已被广泛研究(Tolbert and Essner, 1981; Trelease, 1984; Kindl, 1992)。其中一种存在于绿叶中,在乙醇酸代谢中起重要作用,这与光呼吸(photorespiration)有关,光呼吸是一个消耗氧气并释放二氧化碳的过程。光呼吸涉及过氧化物酶体、线粒体和叶绿体之间的协同相互作用;因此,这三种细胞器通常在空间上彼此紧密关联(图2.19A)。光呼吸的生物学功能仍有待确定(Taiz and Zeiger, 2002)。
第二种类型的过氧化物酶体存在于萌发种子的胚乳或子叶中,在那里它通过一系列称为乙醛酸循环(glyoxylate cycle)的反应在脂肪转化为碳水化合物的过程中起重要作用。因此,这些过氧化物酶体也被称为乙醛酸循环体(glyoxysome)。这两种类型的过氧化物酶体是可以相互转化的(Kindl, 1992; Nishimura et al., 1993, 1998)。例如,在萌发的早期阶段,一些种子的子叶基本上没有光照。随着子叶逐渐暴露在光线下,它们可能会变绿。随着脂肪的消耗和叶绿体的出现,乙醛酸循环体转化为叶型过氧化物酶体。当组织经历衰老时,乙醛酸循环体的特性可能会重新出现。
几项研究表明,植物过氧化物酶体与质体(plastid)和线粒体一样,是依赖于肌动蛋白(actin)运动的细胞器(Collings et al., 2002; Jedd and Chua, 2002; Mano et al., 2002; Mathur et al., 2002)。韭葱(Allium porrum)和拟南芥(Arabidopsis)中的过氧化物酶体已被证明沿着肌动蛋白丝束进行动态运动(Collings et al., 2002; Mano et al., 2002),拟南芥中的过氧化物酶体达到接近10\upmum\cdots-1的峰值速度(Jedd and Chua, 2002)。此外,拟南芥中的过氧化物酶体已被证明是由肌球蛋白(myosin)驱动的(Jedd and Chua, 2002)。
图2.19
甜菜(Beta vulgaris, A)和烟草(Nicotiana tabacum, B)叶细胞中的细胞器。包围过氧化物酶体的单位膜可以与其他细胞器的双层膜包被形成对比。B中的过氧化物酶体含有晶体。在A的叶绿体和B的线粒体中可以看到一些核糖体。(来自Esau, 1977。)
❙ 液泡
与质体和细胞壁(cell wall)的存在一起,液泡是区分植物细胞和动物细胞的三个特征之一。如前所述,液泡是由单层膜即液泡膜(tonoplast)或液泡膜(vacuolar membrane)包围的细胞器(图2.2)。它们是多功能细胞器,在形态、大小、内容和功能动态方面具有广泛的多样性(Wink, 1993; Marty, 1999)。一个细胞可能包含不止一种液泡。一些液泡主要作为储存细胞器,另一些则作为溶解区室(lytic compartments)。这两种类型的液泡可以通过特定的液泡膜内在蛋白(tonoplast integral (intrinsic) proteins, TIPs)的存在来表征:例如,α-TIP与蛋白质储存液泡的液泡膜相关,而γ-TIP则定位于溶解液泡的液泡膜。两种类型的TIP可能共定位于大液泡的同一液泡膜上,这显然是细胞扩大过程中两种液泡合并的结果(Paris et al., 1996; Miller and Anderson, 1999)。
许多分生组织(meristematic)植物细胞含有大量小液泡。随着细胞扩大,液泡增大并融合成一个大的液泡(图2.20)。事实上,细胞大小的增加主要涉及液泡的扩大。在成熟细胞中,液泡可能占据高达90%的体积,其余细胞质(cytoplasm)由紧贴细胞壁的薄外周层组成。通过用"廉价"(就能量而言)的液泡内容物填充如此大比例的细胞,植物不仅节省了"昂贵"的富含氮的细胞质材料,还在富含氮的薄层细胞质和原生质体(protoplast)的外部环境之间获得了大的表面积(Wiebe, 1978)。作为一种选择性通透膜(selectively permeable membrane),液泡膜参与调节与液泡相关的渗透现象(osmotic phenomena)。这种策略的直接结果是组织刚性(tissue rigidity)的发展,这是液泡和液泡膜的主要作用之一。
非蛋白质储存液泡的主要成分是水,其他成分根据植物类型、器官和细胞及其发育和生理状态而有所不同(Nakamura and Matsuoka, 1993; Wink, 1993)。除了无机离子如Ca2+、Cl-、 K+、Na+、NO3-和PO42-外,这些液泡通常还含有糖类、有机酸和氨基酸,其水溶液通常称为细胞液(cell sap)。有时液泡中某种物质的浓度足够大,可以形成晶体。草酸钙(calcium oxalate)晶体可以呈现不同的形式(第3章),尤其...
图2.21
含单宁的液泡在敏感植物(Mimosa pudica)的叶细胞中。电子致密的单宁几乎填满了这个细胞的中央液泡。
在大多数情况下,液泡并不合成它们所积累的分子,而是必须从细胞质的其他部分接收这些分子。代谢物和无机离子通过液泡膜(tonoplast)的运输受到严格控制,以确保细胞的最佳功能(Martinoia, 1992; Nakamura and Matsuoka, 1993; Wink, 1993)。
液泡是各种代谢物的重要储存区。初级代谢物(primary metabolites)——在细胞代谢中起基本作用的物质——如糖和有机酸,仅在液泡中暂时储存。例如,在许多物种的光合作用叶片中,白天产生的大部分糖储存在叶肉细胞的液泡中,然后在夜间从液泡中移出,输送到植物的其他部分。在CAM植物中,苹果酸在夜间储存在液泡中,白天从液泡中释放并脱羧,CO2随后在叶绿体中被卡尔文循环同化(Kluge et al., 1982; Smith, 1987)。在种子中,液泡是蛋白质储存的主要场所(HermaNand Larkins, 1999)。
液泡还从细胞质的其他部分隔离有毒的次级代谢物(secondary metabolites),如尼古丁(一种生物碱)和单宁(酚类化合物)(图2.21)。次级代谢物在植物的初级代谢中不起明显作用。这些物质可能永久地隔离在液泡中。液泡中积累的大量次级代谢物不仅对植物本身有毒,而且对病原体、寄生虫和/或草食动物也有毒,因此它们在植物防御中起着重要作用。储存在液泡中的一些次级代谢物是无毒的,但在液泡破裂时通过水解转化为剧毒衍生物,如氰化物、芥子油和苷元(Matile, 1982; Boller and Wiemken, 1986)。因此,细胞质的解毒和防御性化学物质的储存可以被视为液泡的附加功能。
液泡通常是色素沉积的场所。植物细胞的蓝色、紫色、深红色和猩红色通常是由一组称为花青素(anthocyanins)的色素引起的。这些色素通常局限于表皮细胞。与大多数其他植物色素(如叶绿素、类胡萝卜素)不同,花青素易溶于水,并以溶液形式存在于液泡中。它们负责许多水果(葡萄、李子、樱桃)和蔬菜(萝卜、芜菁、卷心菜)以及大量花卉(天竺葵、飞燕草、玫瑰、矮牵牛、牡丹)的红色和蓝色,并可能用于吸引动物进行授粉和种子传播。花青素被认为与Brassica幼苗外围细胞层液泡中钼的隔离有关(Hale et al., 2001)。在少数植物科中,另一类水溶性色素——含氮的甜菜碱(betalains)——负责一些黄色和红色。这些植物都属于藜目(Chenopodiales),缺乏花青素。甜菜和九重葛花的红色是由于甜菜红素(红色甜菜碱)的存在。黄色甜菜碱称为甜菜黄素(Piattelli, 1981)。
花青素(anthocyanins)也是导致一些树叶在秋季呈现鲜艳红色的原因。这些色素在寒冷、阳光充足的天气条件下形成,此时叶子停止产生叶绿素(chlorophyll)。随着现有叶绿素的分解,新形成的花青素得以显现。在不形成花青素色素的叶子中,秋季叶绿素的分解可能会使叶绿体中已经存在的更稳定的黄色到橙色类胡萝卜素(carotenoid)色素显现出来。最壮观的秋季色彩出现在秋季凉爽、晴朗的年份(Kozlowski和Pallardy, 1997)。
叶子中的花青素起什么作用?在红枝山茱萸(Cornus stolonifera)中,花青素在秋季在栅栏叶肉层(palisade mesophyll layer)形成色素层,在叶子脱落前减少叶绿体对光的捕获。有人认为,花青素对叶绿素的这种光学遮蔽降低了叶片细胞衰老时发生光氧化损伤(photo-oxidative damage)的风险,否则这种损伤可能会降低从衰老叶片中回收养分的效率(Feild等, 2001)。除了保护叶片免受光氧化损伤外,有证据表明花青素还能防止光抑制(photoinhibition)(Havaux和Kloppstech, 2001; Lee, D. W.和Gould, 2002; Steyn等, 2002),即到达光系统II反应中心的过量激发导致的光合效率下降。光抑制在下层植物中很常见,当它们突然暴露在穿过上层冠层瞬时开口(树叶在微风中摇曳时形成)的斑驳阳光(sunflecks)下时就会发生(Pearcy, 1990)。
作为溶酶体区室(lytic compartments),液泡(vacuoles)参与大分子的分解和细胞内成分的循环利用。整个细胞器,如衰老的质体和线粒体(mitochondria),可能会被含有大量水解酶(hydrolytic enzymes)和氧化酶(oxidizing enzymes)的液泡吞噬并随后降解。大型中央液泡可以隔离水解酶(hydrolases),当液泡膜(tonoplast)破裂时,可能导致细胞质(cytoplasm)完全自溶(autolysis),如在分化中的管状元件(tracheary elements)的程序性细胞死亡(programmed cell death)期间(第10章)。由于这种消化活性,所谓的溶酶体液泡(lytic vacuoles)在功能上可与动物细胞中称为溶酶体(lysosomes)的细胞器相媲美。
长期以来,人们一直认为新的液泡是由光滑内质网(smooth ER)的特殊区域扩张或由高尔基体(Golgi apparatus)衍生的囊泡(vesicles)形成的。大多数证据支持液泡从内质网从头形成(de novo formation)(Robinson, 1985; Hörtensteiner等, 1992; Herman等, 1994)。
❙ 核糖体
核糖体是直径仅约17至23纳米的小颗粒(图2.22),由蛋白质和RNA组成(Davies和Larkins, 1980)。尽管核糖体中的蛋白质分子数量远远超过RNA分子数量,但RNA约占核糖体质量的60%。它们是氨基酸连接在一起形成蛋白质的场所,在代谢活跃细胞的细胞质中含量丰富(Lake, 1981)。每个核糖体由两个亚基组成,一个较小,另一个较大,由特定的核糖体RNA和蛋白质分子构成。核糖体既自由存在于细胞质中,也附着在内质网和核膜的外表面。它们是迄今为止数量最多的细胞结构,也存在于细胞核、质体和线粒体中。如前所述,质体和线粒体的核糖体大小与细菌的相似。
积极参与蛋白质合成的核糖体以称为多核糖体(polysomes)或多聚核糖体(polyribosomes)(图2.22)的簇或聚集体形式存在,由携带来自细胞核的遗传信息的信使RNA分子连接在一起。用于合成蛋白质的氨基酸由位于细胞质中的转运RNA带到多核糖体。蛋白质的合成,称为翻译(translation),消耗的能量比任何其他生物合成过程都多。这些能量由三磷酸鸟苷(GTP)的水解提供。
图2.22
核糖体。A,玉米(Zea mays)叶片维管束鞘细胞中的核糖体。箭头指向一束肌动蛋白丝。B,烟草(Nicotiana tabacum)叶细胞中附着在内质网表面的多核糖体(多聚核糖体)。(B,来自Esau, 1977。)
由核基因编码的多肽(蛋白质)的合成在位于细胞质中的多核糖体上启动,并遵循两条不同的途径之一。(1)那些参与合成注定要进入内质网的多肽的多核糖体在翻译过程的早期就与内质网相关联。多肽及其相关的多核糖体通过位于每个多肽氨基端的靶向信号或信号肽(signal peptide)被引导到内质网。随着多肽合成的继续,多肽被转移穿过膜进入内质网腔(或插入其中,在整合蛋白的情况下)。(2)那些参与合成注定要进入细胞质或导入细胞核、线粒体、质体或过氧化物酶体的多肽的多核糖体仍然自由存在于细胞质中。从自由多核糖体释放的多肽要么留在细胞质中,要么通过靶向序列(targeting sequence)被引导到适当的细胞组分(Holtzman, 1992)。膜结合和自由核糖体在结构和功能上都是相同的,仅在它们在任何给定时间制造的蛋白质上有所不同。第一章 植物体的结构和发育——概览